

Structure and Dynamics of Membrane Proteins from Solid-State NMR
- PMID: 29498890
- PMCID: PMC6312106
- DOI: 10.1146/annurev-biophys-070816-033712
Structure and Dynamics of Membrane Proteins from Solid-State NMR
Abstract
Solid-state nuclear magnetic resonance (SSNMR) spectroscopy elucidates membrane protein structures and dynamics in atomic detail to yield mechanistic insights. By interrogating membrane proteins in phospholipid bilayers that closely resemble biological membranes, SSNMR spectroscopists have revealed ion conduction mechanisms, substrate transport dynamics, and oligomeric interfaces of seven-transmembrane helix proteins. Research has also identified conformational plasticity underlying virus-cell membrane fusions by complex protein machineries, and β-sheet folding and assembly by amyloidogenic proteins bound to lipid membranes. These studies collectively show that membrane proteins exhibit extensive structural plasticity to carry out their functions. Because of the inherent dependence of NMR frequencies on molecular orientations and the sensitivity of NMR frequencies to dynamical processes on timescales from nanoseconds to seconds, SSNMR spectroscopy is ideally suited to elucidate such structural plasticity, local and global conformational dynamics, protein-lipid and protein-ligand interactions, and protonation states of polar residues. New sensitivity-enhancement techniques, resolution enhancement by ultrahigh magnetic fields, and the advent of 3D and 4D correlation NMR techniques are increasingly aiding these mechanistically important structural studies.
Keywords: conformational dynamics; ion channels; magic-angle-spinning NMR; transporters; viral fusion proteins.
Figures
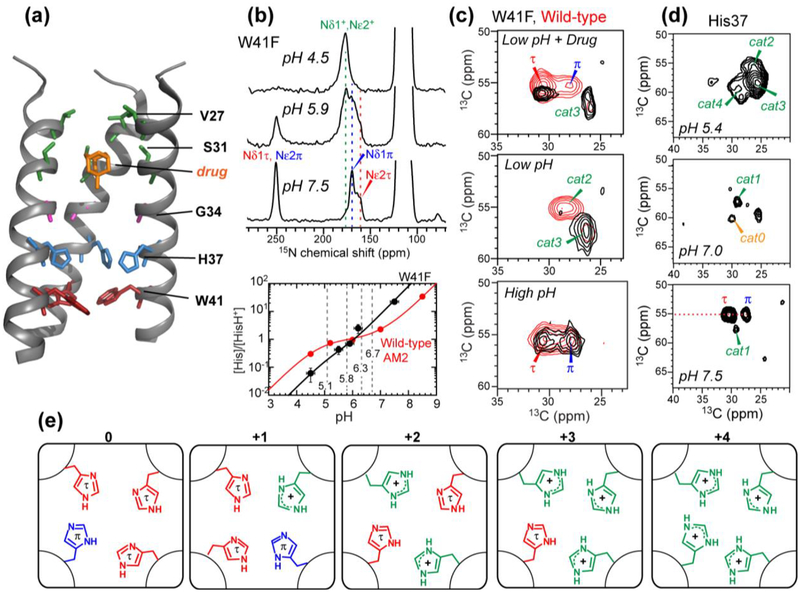
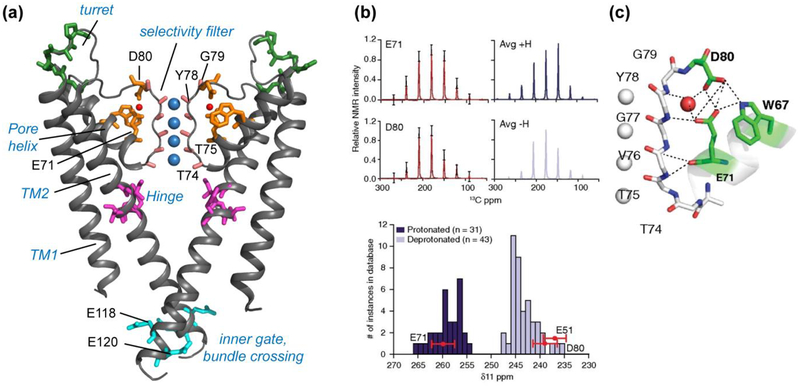
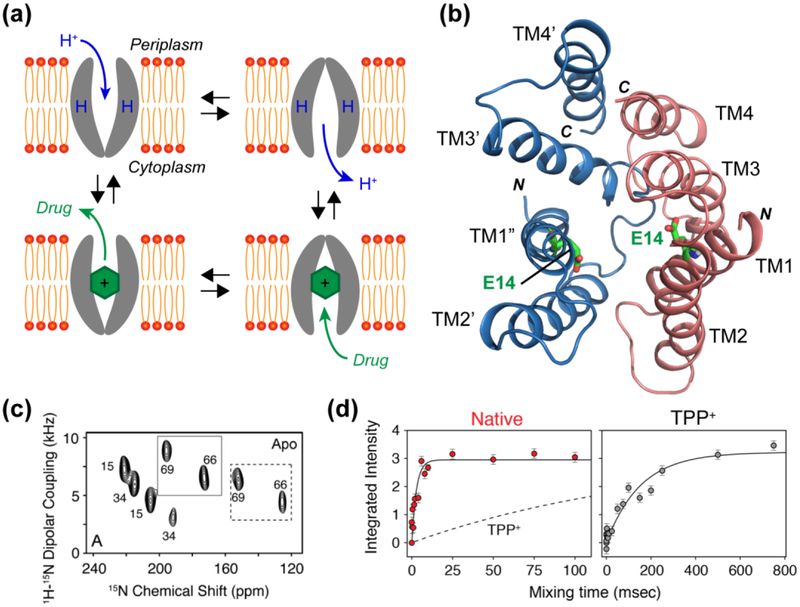
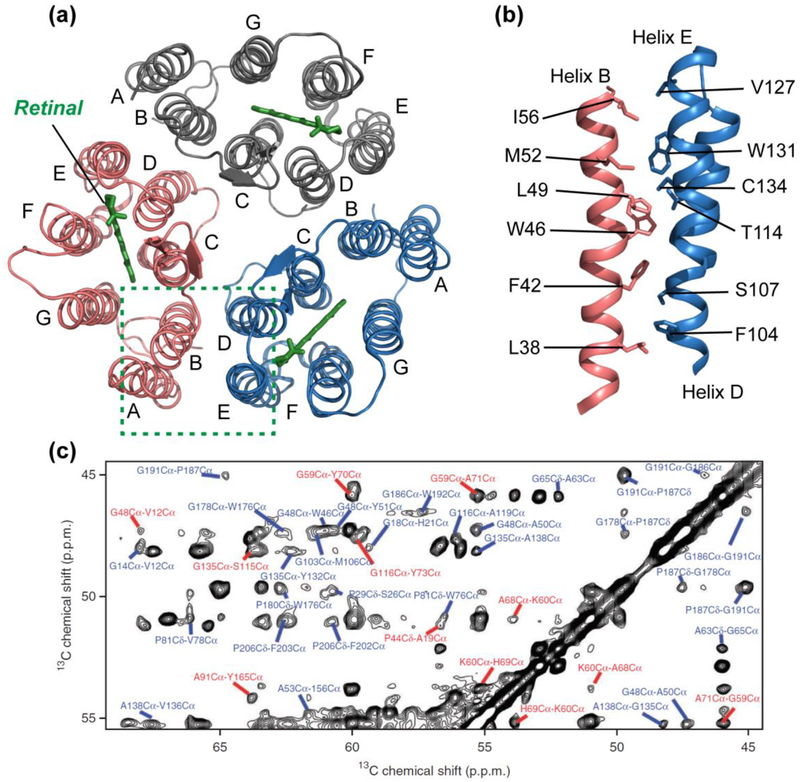
References
-
- Akinlolu R, Nam M, Qiang W. 2015. Competition between Fibrillation and Induction of Vesicle Fusion for the Membrane-Associated 40-Residue b-Amyloid Peptides, Biochemistry 54:3416–3419 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
