

Multidimensional chemobehavior analysis of flavonoids and neuroactive compounds in zebrafish
- PMID: 29499247
- PMCID: PMC5912343
- DOI: 10.1016/j.taap.2018.02.019
Multidimensional chemobehavior analysis of flavonoids and neuroactive compounds in zebrafish
Abstract
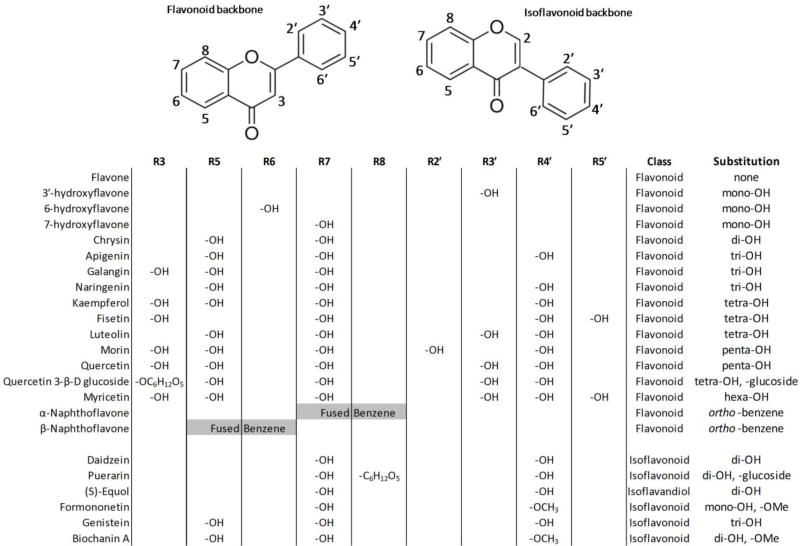
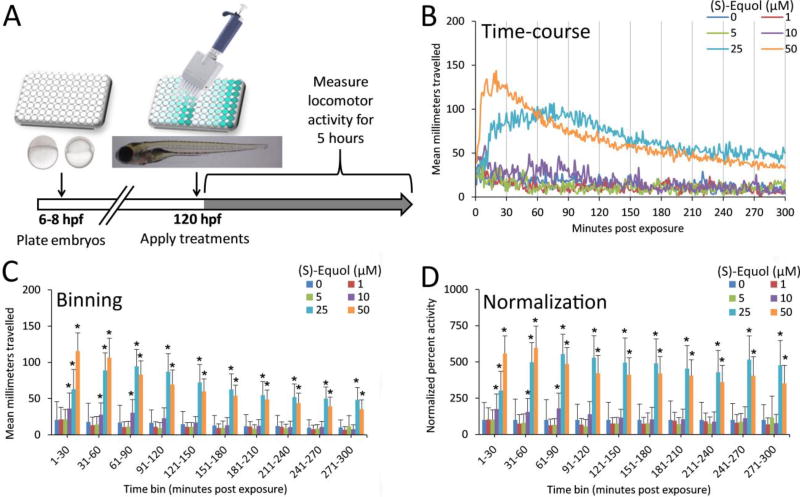
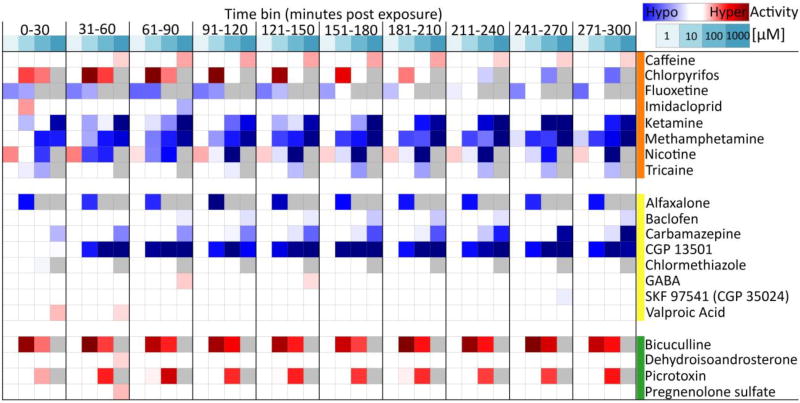
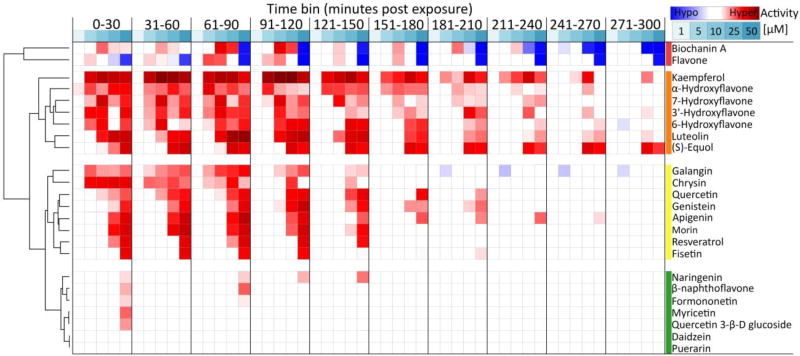
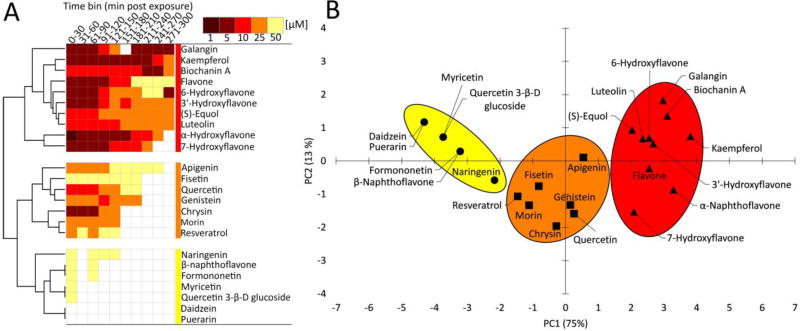
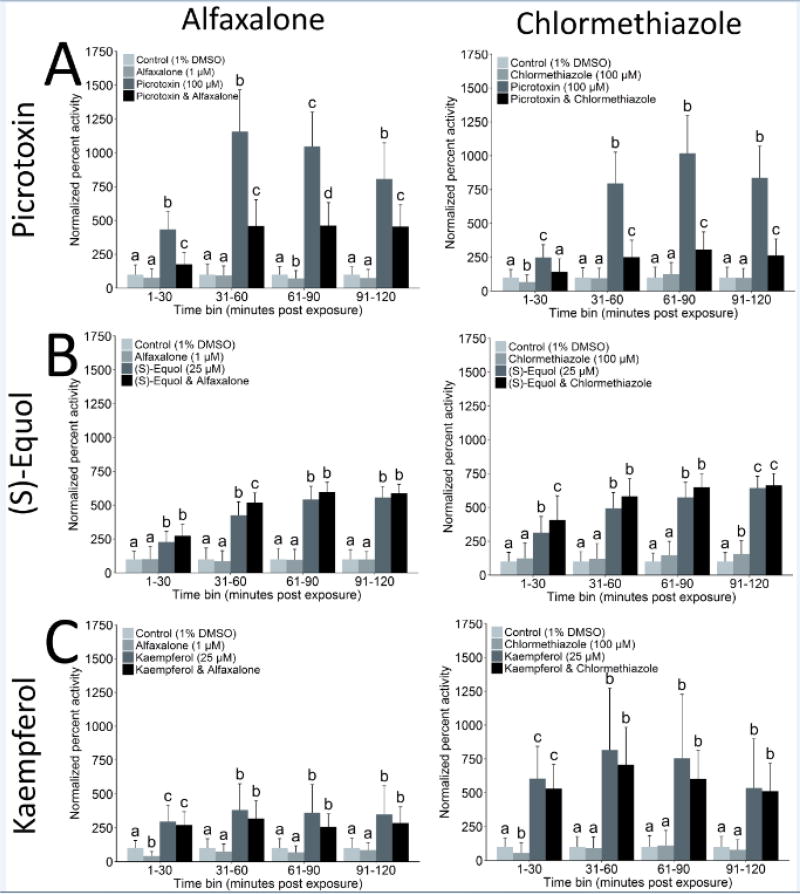
The comparative analysis of complex behavioral phenotypes is valuable as a reductionist tool for both drug discovery and defining chemical bioactivity. Flavonoids are a diverse class of chemicals that elicit robust neuroactive and hormonal actions, though bioactivity information is limited for many, particularly for neurobehavioral endpoints. Here, we used a zebrafish larval chemomotor response (LCR) bioassay to comparatively evaluate a suite of 24 flavonoids, and in addition a panel of 30 model neuroactive compounds representing diverse modes of action (e.g. caffeine, chlorpyrifos, methamphetamine, nicotine, picrotoxin). Naïve larval zebrafish were exposed to concentration ranges of each compound at 120 hour post-fertilization (hpf) and locomotor activity measured for 5 h. The model neuroactive compounds were largely behaviorally bioactive (20 of 30) with most effects phenotypic of their known modes of action. Flavonoids rapidly and broadly elicited hyperactive locomotor effects (22 of 24). Multidimensional analyses compared responses over time and identified three distinct bioactive groups of flavonoids based on efficacy and potency. Using GABAergics to modulate hyperactive responses, two flavonoids, (S)-equol and kaempferol were tested for GABAA receptor antagonism, as well as a known GABAA receptor antagonist, picrotoxin. Pharmacological intervention with positive allosteric modulators of the GABAA receptor, alfaxalone and chlormethiazole, ameliorated the hyperactive response to picrotoxin, but not for (S)-equol or kaempferol. Taken together, these studies demonstrate that flavonoids are differentially bioactive and that the chemobehavioral effects likely do not involve a GABAA receptor mediated mode of action. Overall, the integrative zebrafish platform provides a useful framework for comparatively evaluating high-content chemobehavioral data for sets of structurally- and mechanistically-related flavonoids and neuroactive compounds.
Keywords: Behavior; Flavonoids; GABA receptor; Locomotion; Neurotoxicity; Zebrafish.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare they have no potential conflicts of interest.
Figures
References
-
- Afrikanova T, Serruys AS, Buenafe OE, Clinckers R, Smolders I, de Witte PA, Crawford AD, Esguerra CV. Validation of the zebrafish pentylenetetrazol seizure model: locomotor versus electrographic responses to antiepileptic drugs. PloS one. 2013;8:e54166. doi: 10.1371/journal.pone.0054166. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
