

Functional Anti-TIGIT Antibodies Regulate Development of Autoimmunity and Antitumor Immunity
- PMID: 29500245
- PMCID: PMC5893394
- DOI: 10.4049/jimmunol.1700407
Functional Anti-TIGIT Antibodies Regulate Development of Autoimmunity and Antitumor Immunity
Abstract
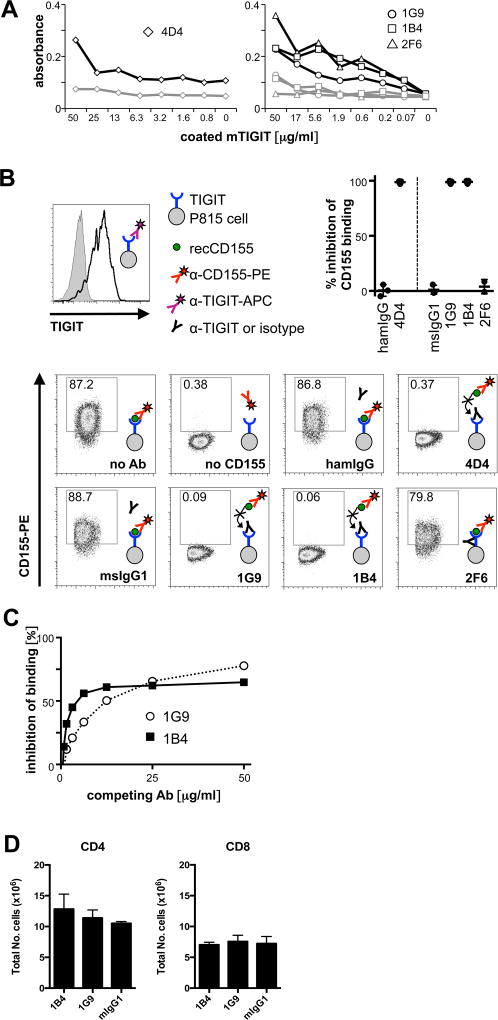
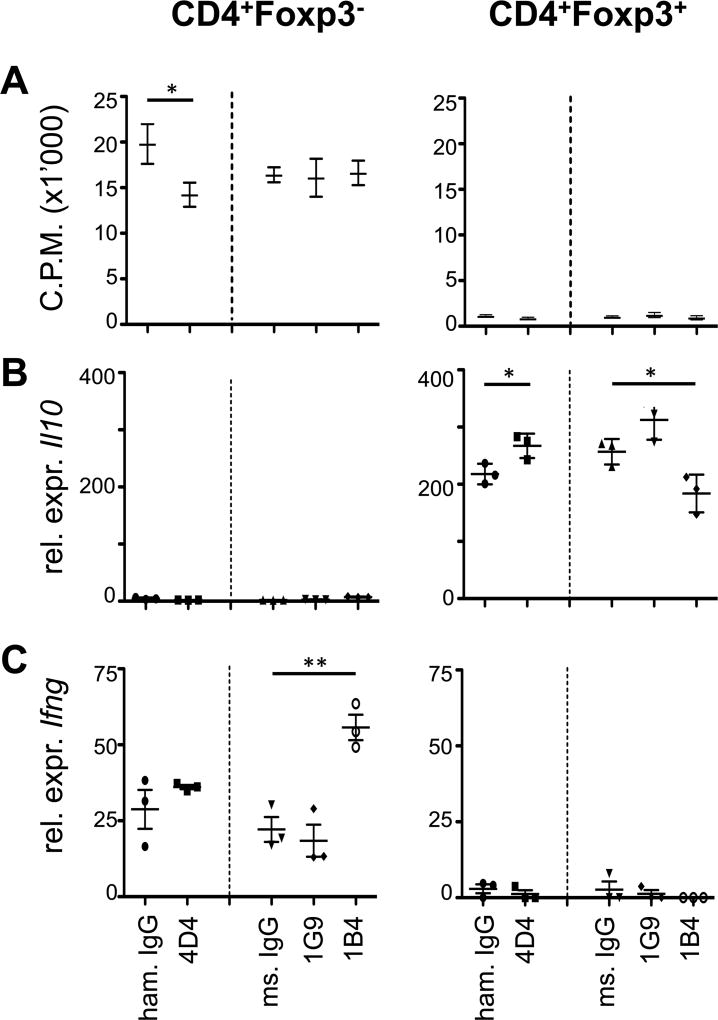
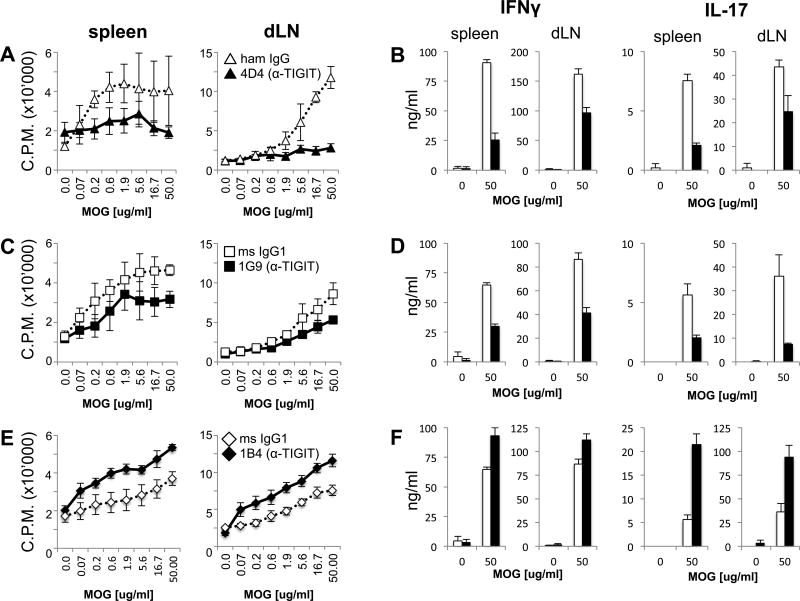
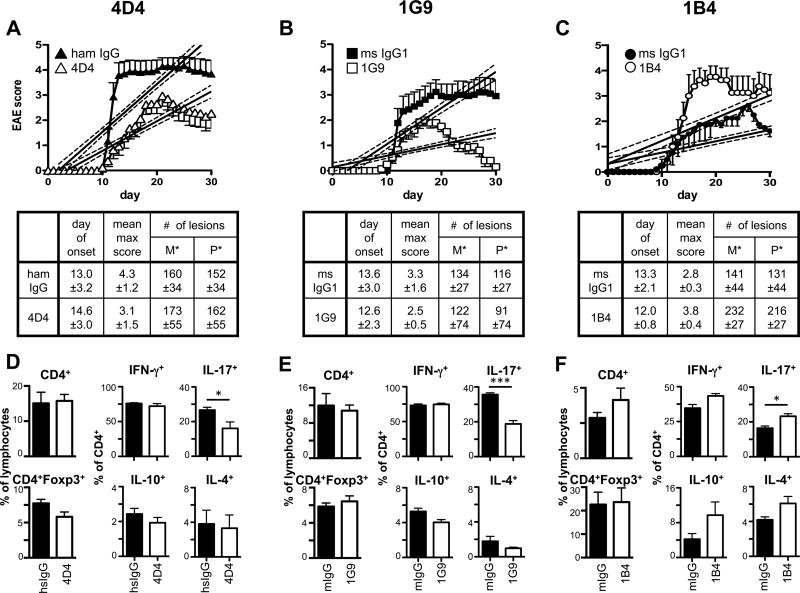
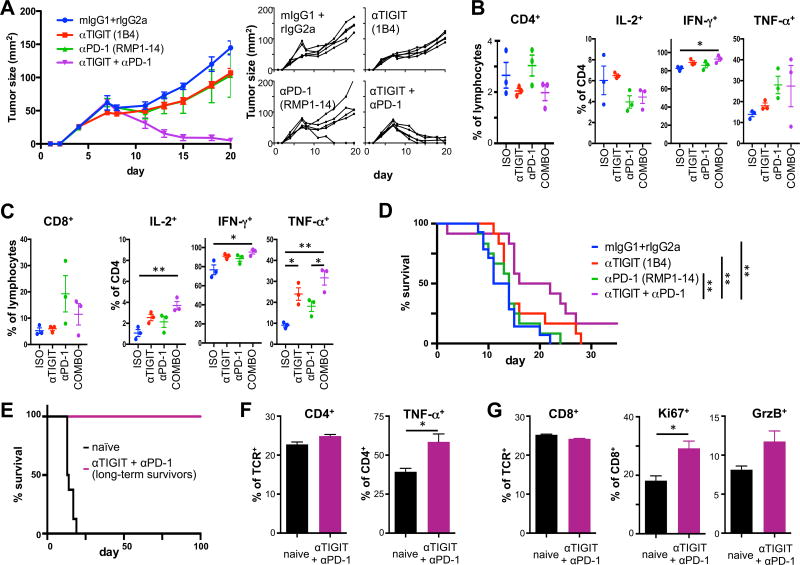
Coinhibitory receptors, such as CTLA-4 and PD-1, play a critical role in maintaining immune homeostasis by dampening T cell responses. Recently, they have gained attention as therapeutic targets in chronic disease settings where their dysregulated expression contributes to suppressed immune responses. The novel coinhibitory receptor TIGIT (T cell Ig and ITIM domain) has been shown to play an important role in modulating immune responses in the context of autoimmunity and cancer. However, the molecular mechanisms by which TIGIT modulates immune responses are still insufficiently understood. We have generated a panel of monoclonal anti-mouse TIGIT Abs that show functional properties in mice in vivo and can serve as important tools to study the underlying mechanisms of TIGIT function. We have identified agonistic as well as blocking anti-TIGIT Ab clones that are capable of modulating T cell responses in vivo. Administration of either agonist or blocking anti-TIGIT Abs modulated autoimmune disease severity whereas administration of blocking anti-TIGIT Abs synergized with anti-PD-1 Abs to affect partial or even complete tumor regression. The Abs presented in this study can thus serve as important tools for detailed analysis of TIGIT function in different disease settings and the knowledge gained will provide valuable insight for the development of novel therapeutic approaches targeting TIGIT.
Copyright © 2018 by The American Association of Immunologists, Inc.
Conflict of interest statement
A.C.A. is a member of the SAB for Potenza Therapeutics, Tizona Therapeutics, and Idera Pharmaceuticals, which have interests in cancer immunotherapy. V.K.K. has an ownership interest and is a member of the SAB for Potenza Therapeutics and Tizona Therapeutics. A.C.A.’s and V.K.K.’s interests were reviewed and managed by the Brigham and Women’s Hospital and Partners Healthcare in accordance with their conflict of interest policies. T. Kondo is an employee of Mitsubishi Tanabe Pharma Corporation and was supported by its Scholarship Program. R.K.J. received consultant fees from Merck, Ophthotech, Pfizer, SPARC, SynDevRx, XTuit; owns equity in Enlight, Ophthotech, SynDevRx, XTuit; and serves on the Board of Directors of XTuit and the Boards of Trustees of Tekla Healthcare Investors, Tekla Life Sciences Investors, Tekla Healthcare Opportunities Fund, Tekla World Healthcare Fund. Neither any reagent nor any funding from these organizations was used in this study.
Figures
References
-
- Joller N, Lozano E, Burkett PR, Patel B, Xiao S, Zhu C, Xia J, Tan TG, Sefik E, Yajnik V, Sharpe AH, Quintana FJ, Mathis D, Benoist C, Hafler DA, Kuchroo VK. Treg Cells Expressing the Coinhibitory Molecule TIGIT Selectively Inhibit Proinflammatory Th1 and Th17 Cell Responses. Immunity. 2014;40:569–581. - PMC - PubMed
-
- Yu X, Harden K, Gonzalez LC, Francesco M, Chiang E, Irving B, Tom I, Ivelja S, Refino CJ, Clark H, Eaton D, Grogan JL. The surface protein TIGIT suppresses T cell activation by promoting the generation of mature immunoregulatory dendritic cells. Nat Immunol. 2009;10:48–57. - PubMed
-
- Johnston RJ, Comps-Agrar L, Hackney J, Yu X, Huseni M, Yang Y, Park S, Javinal V, Chiu H, Irving B, Eaton DL, Grogan JL. The Immunoreceptor TIGIT Regulates Antitumor and Antiviral CD8(+) T Cell Effector Function. Cancer cell. 2014;26:923–937. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
