

Snapshots of C-S Cleavage in Egt2 Reveals Substrate Specificity and Reaction Mechanism
- PMID: 29503207
- PMCID: PMC5959753
- DOI: 10.1016/j.chembiol.2018.02.002
Snapshots of C-S Cleavage in Egt2 Reveals Substrate Specificity and Reaction Mechanism
Abstract
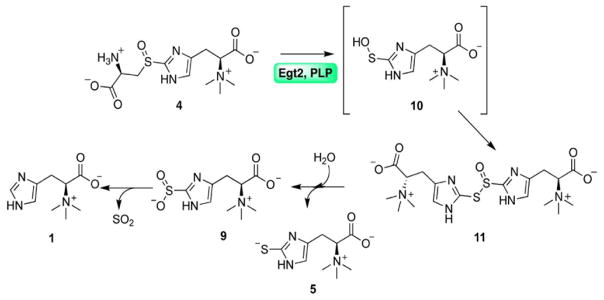
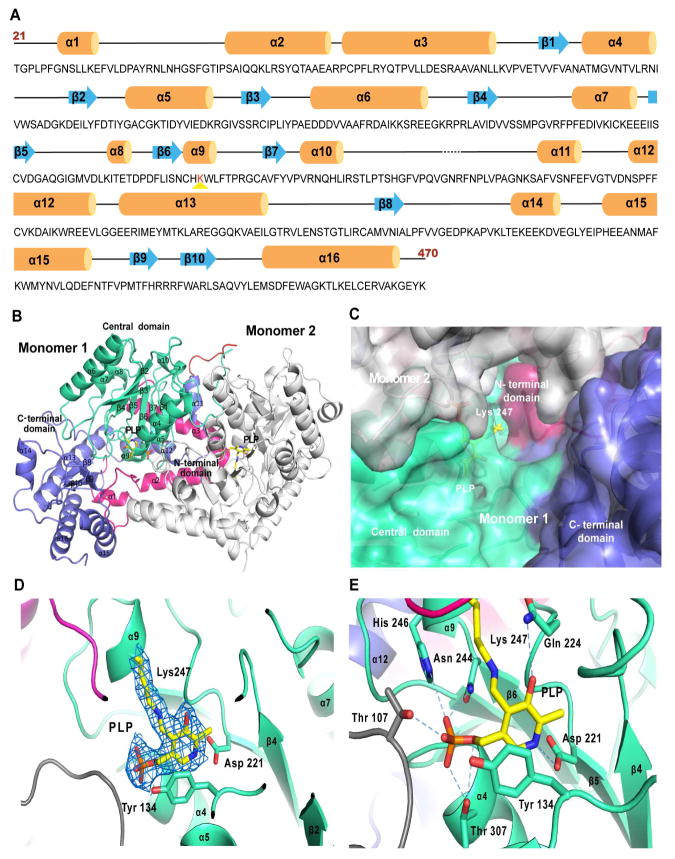
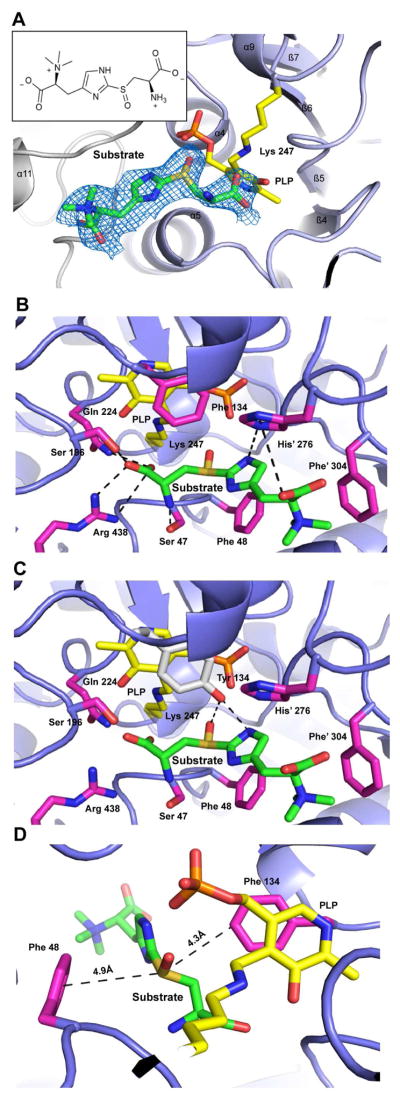
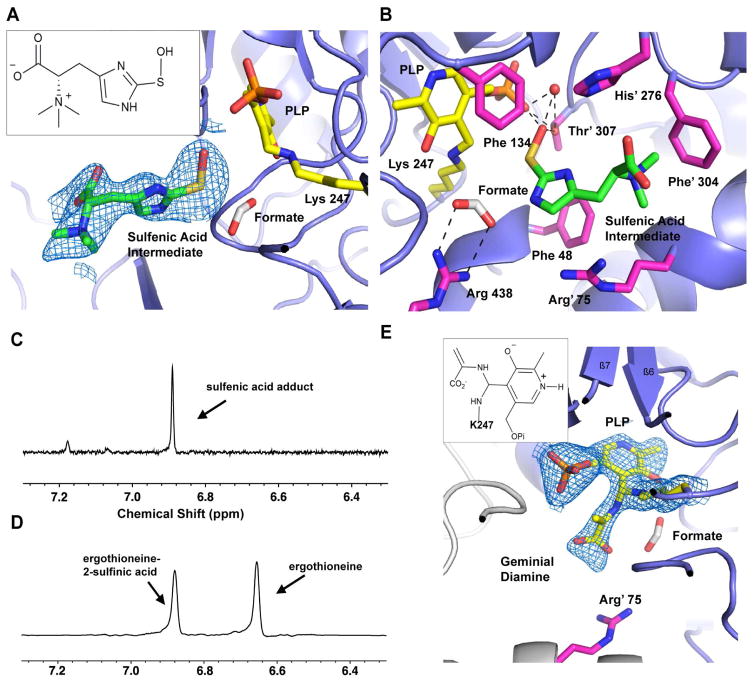
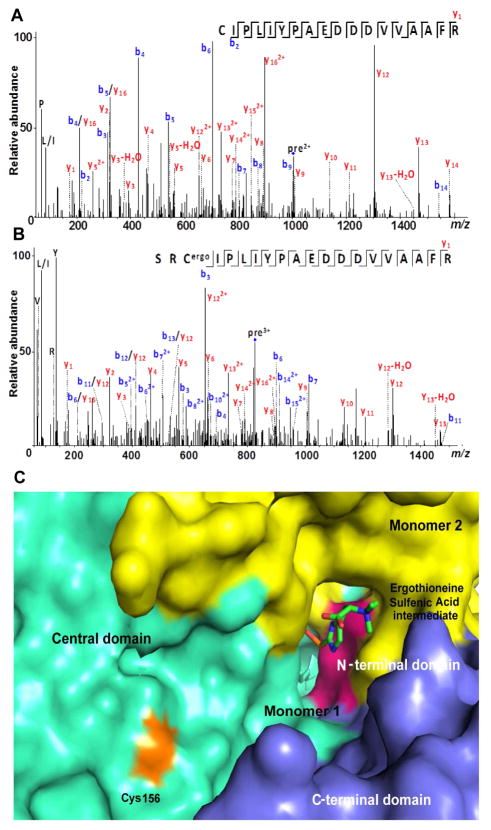
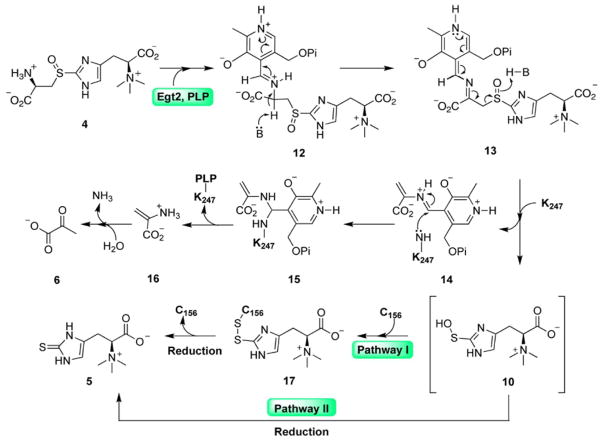
Sulfur incorporation in the biosynthesis of ergothioneine, a histidine thiol derivative, differs from other well-characterized transsulfurations. A combination of a mononuclear non-heme iron enzyme-catalyzed oxidative C-S bond formation and a subsequent pyridoxal 5'-phosphate (PLP)-mediated C-S lyase reaction leads to the net transfer of a sulfur atom from a cysteine to a histidine. In this study, we structurally and mechanistically characterized a PLP-dependent C-S lyase Egt2, which mediates the sulfoxide C-S bond cleavage in ergothioneine biosynthesis. A cation-π interaction between substrate and enzyme accounts for Egt2's preference of sulfoxide over thioether as a substrate. Using mutagenesis and structural biology, we captured three distinct states of the Egt2 C-S lyase reaction cycle, including a labile sulfenic intermediate captured in Egt2 crystals. Chemical trapping and high-resolution mass spectrometry were used to confirm the involvement of the sulfenic acid intermediate in Egt2 catalysis.
Keywords: C-S bond cleavage; X-ray crystallography; biosynthetic pathway; chemical trapping; enzymology; ergothioneine.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Conflict of interest statement
The authors declare competing financial interests. A patent (WO 2014100752 A1) on ergothioneine production through metabolic engineering has been submitted.
Figures
References
-
- Allegrini A, Astegno A, La Verde V, Dominici P. Characterization of C-S lyase from Lactobacillus delbrueckii subsp bulgaricus ATCC BAA-365 and its potential role in food flavour applications. J Biochem. 2017;161:349–360. - PubMed
-
- Allison WS. Formation and reactions of sulfenic acids in proteins. Acc Chem Res. 1976;9:293–299.
-
- Aruoma OI, Spencer JPE, Mahmood N. Protection Against Oxidative Damage and Cell Death by the Natural Antioxidant Ergothioneine. Food Chem Toxicol. 1999;37:1043–1053. - PubMed
-
- Baldwin JE, Bradley M. Isopenicillin N synthase: mechanistic studies. Chem Rev. 1990;90:1079–1088.
-
- Bartholomeus Kuettner E, Hilgenfeld R, Weiss MS. Purification, characterization, and crystallization of alliinase from garlic. Arch Biochem Biophys. 2002;402:192–200. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
