

Heligmosomoides polygyrus Venom Allergen-like Protein-4 (HpVAL-4) is a sterol binding protein
- PMID: 29505764
- PMCID: PMC5893428
- DOI: 10.1016/j.ijpara.2018.01.002
Heligmosomoides polygyrus Venom Allergen-like Protein-4 (HpVAL-4) is a sterol binding protein
Abstract
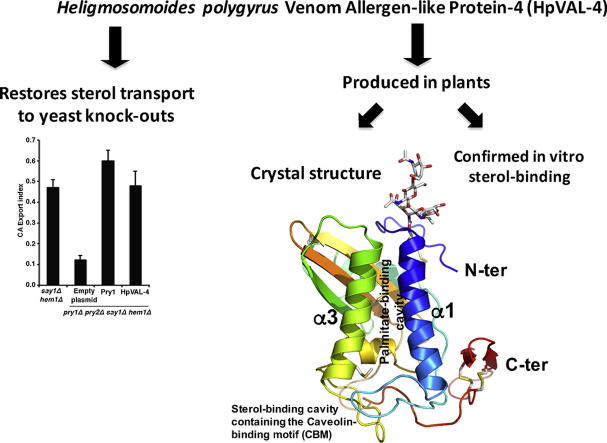
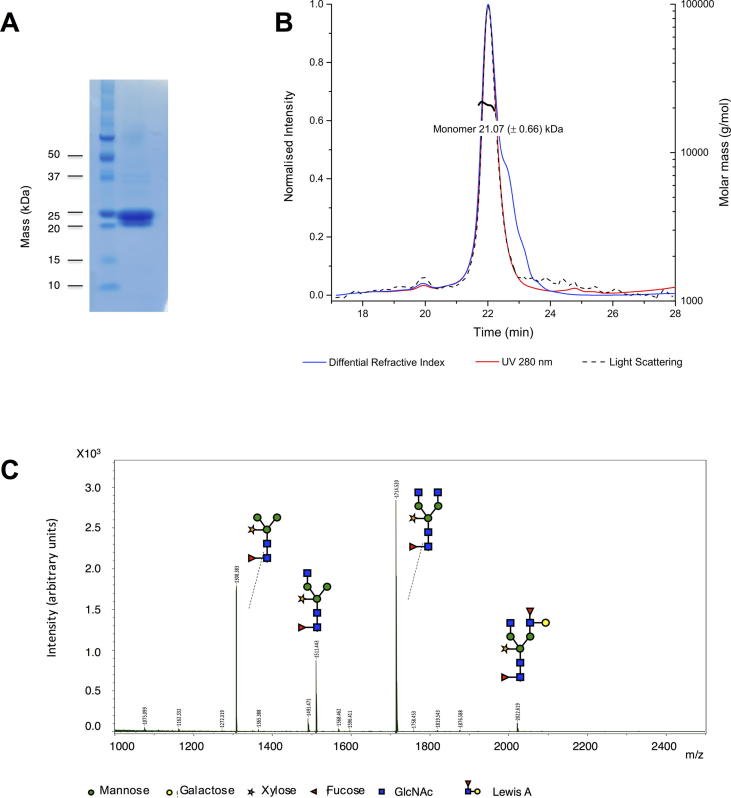
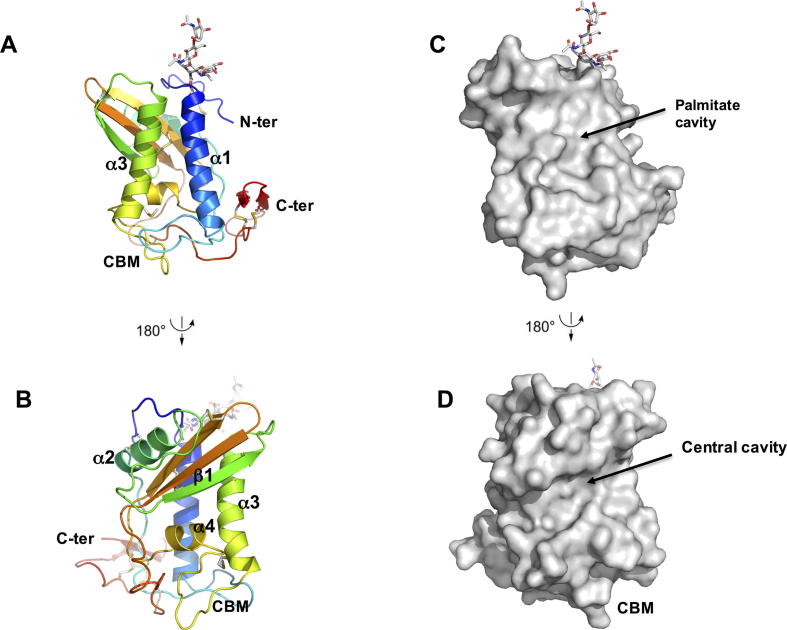
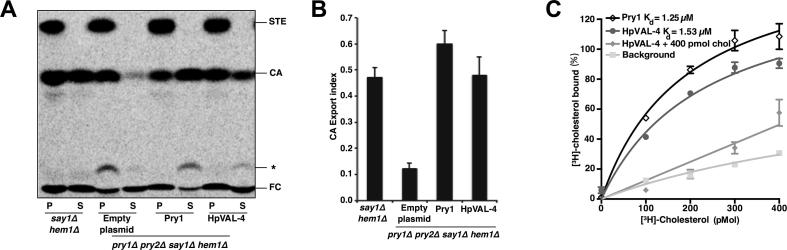
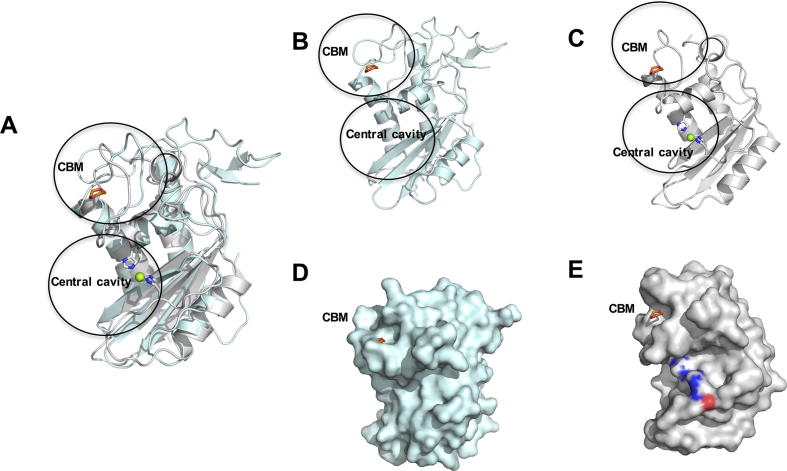
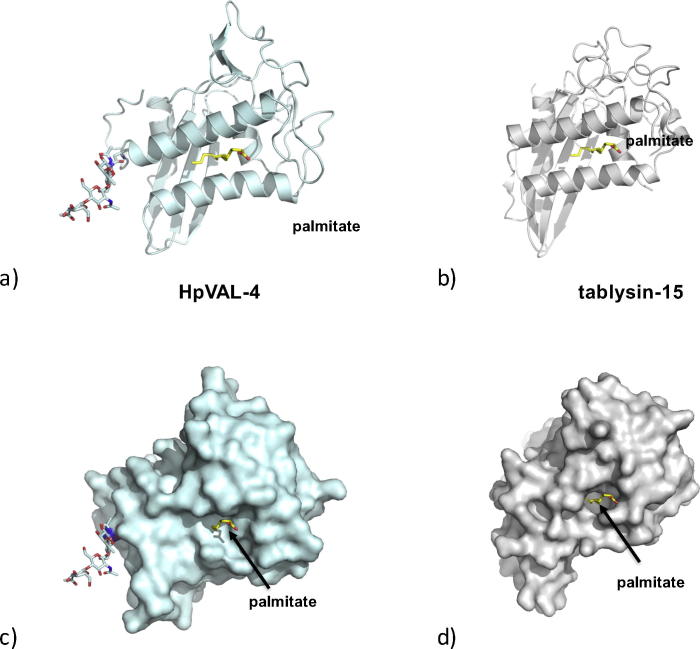
Heligmosomoides polygyrus bakeri is a model parasitic hookworm used to study animal and human helminth diseases. During infection, the parasite releases excretory/secretory products that modulate the immune system of the host. The most abundant protein family in excretory/secretory products comprises the venom allergen-like proteins (VALs), which are members of the SCP/TAPS (sperm-coating protein/Tpx/antigen 5/pathogenesis related-1/Sc7) superfamily. There are >30 secreted Heligmosomoides polygyrus VAL proteins (HpVALs) and these proteins are characterised by having either one or two 15 kDa CAP (cysteine-rich secretory protein (CRISP)/antigen 5/pathogenesis related-1) domains. The first known HpVAL structure, HpVAL-4, refined to 1.9 Å is reported. HpVAL-4 was produced as a homogeneously glycosylated protein in leaves of Nicotiana benthamiana infiltrated with recombinant plasmids, making this plant expression platform amenable for the production of biological products. The overall topology of HpVAL-4 is a three layered αβα sandwich between a short N-terminal loop and a C-terminal cysteine rich extension. The C-terminal cysteine rich extension has two strands stabilized by two disulfide bonds and superposes well with the previously reported extension from the human hookworm Necator americanus Ancylostoma secreted protein-2 (Na-ASP-2). The N-terminal loop is connected to alpha helix 2 via a disulfide bond previously observed in Na-ASP-2. HpVAL-4 has a central cavity that is more similar to the N-terminal CAP domain of the two CAP Na-ASP-1 from Necator americanus. Unlike Na-ASP-2, mammalian CRISP, and the C-terminal CAP domain of Na-ASP-1, the large central cavity of HpVAL-4 lacks the two histidines required to coordinate divalent cations. HpVAL-4 has both palmitate-binding and sterol-binding cavities and is able to complement the in vivo sterol export phenotype of yeast mutants lacking their endogenous CAP proteins. More studies are required to determine endogenous binding partners of HpVAL-4 and unravel the possible impact of sterol binding on immune-modulatory functions.
Keywords: Cysteine-rich secretory protein (CRISP); Excretory–secretory products; Pathogenesis related-1 (PR-1); Sperm coating protein (SCP); Sterol binding; Testis specific proteins (Tpx); Venom antigen 5.
Copyright © 2018 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures

References
-
- Adams P.D., Afonine P.V., Bunkoczi G., Chen V.B., Davis I.W., Echols N., Headd J.J., Hung L.W., Kapral G.J., Grosse-Kunstleve R.W., McCoy A.J., Moriarty N.W., Oeffner R., Read R.J., Richardson D.C., Richardson J.S., Terwilliger T.C., Zwart P.H. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D. 2010;66:213–221. - PMC - PubMed
-
- Asojo O.A., Goud G., Dhar K., Loukas A., Zhan B., Deumic V., Liu S., Borgstahl G.E., Hotez P.J. X-ray structure of Na-ASP-2, a pathogenesis-related-1 protein from the nematode parasite, Necator americanus, and a vaccine antigen for human hookworm infection. J. Mol. Biol. 2005;346:801–814. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
