

N-terminal arginylation generates a bimodal degron that modulates autophagic proteolysis
- PMID: 29507222
- PMCID: PMC5866579
- DOI: 10.1073/pnas.1719110115
N-terminal arginylation generates a bimodal degron that modulates autophagic proteolysis
Abstract
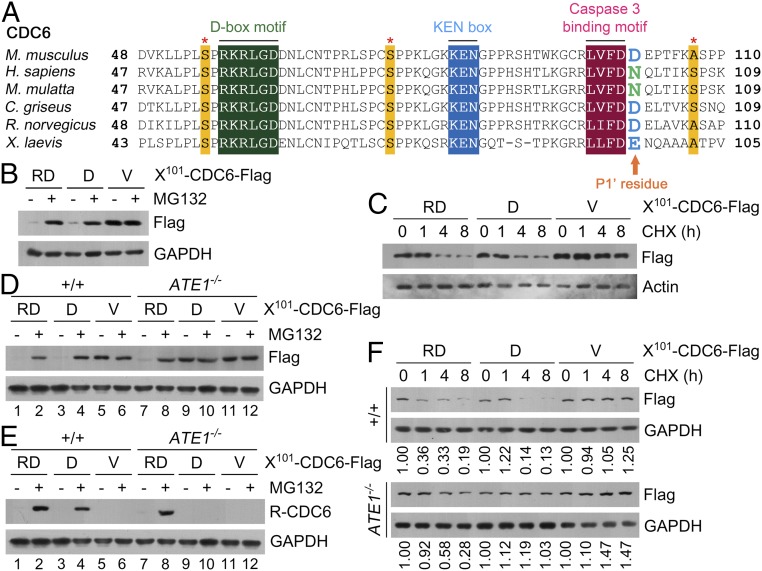
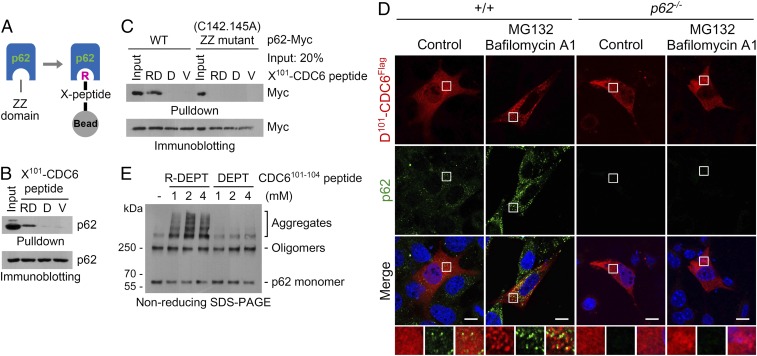
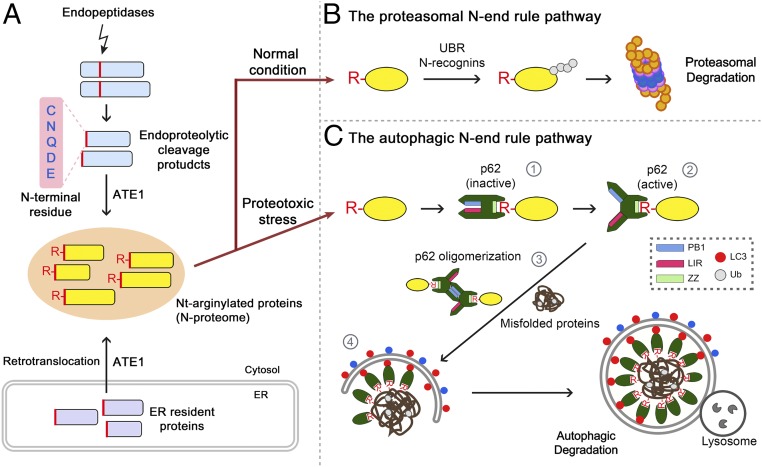
The conjugation of amino acids to the protein N termini is universally observed in eukaryotes and prokaryotes, yet its functions remain poorly understood. In eukaryotes, the amino acid l-arginine (l-Arg) is conjugated to N-terminal Asp (Nt-Asp), Glu, Gln, Asn, and Cys, directly or associated with posttranslational modifications. Following Nt-arginylation, the Nt-Arg is recognized by UBR boxes of N-recognins such as UBR1, UBR2, UBR4/p600, and UBR5/EDD, leading to substrate ubiquitination and proteasomal degradation via the N-end rule pathway. It has been a mystery, however, why studies for the past five decades identified only a handful of Nt-arginylated substrates in mammals, although five of 20 principal amino acids are eligible for arginylation. Here, we show that the Nt-Arg functions as a bimodal degron that directs substrates to either the ubiquitin (Ub)-proteasome system (UPS) or macroautophagy depending on physiological states. In normal conditions, the arginylated forms of proteolytic cleavage products, D101-CDC6 and D1156-BRCA1, are targeted to UBR box-containing N-recognins and degraded by the proteasome. However, when proteostasis by the UPS is perturbed, their Nt-Arg redirects these otherwise cellular wastes to macroautophagy through its binding to the ZZ domain of the autophagic adaptor p62/STQSM/Sequestosome-1. Upon binding to the Nt-Arg, p62 acts as an autophagic N-recognin that undergoes self-polymerization, facilitating cargo collection and lysosomal degradation of p62-cargo complexes. A chemical mimic of Nt-Arg redirects Ub-conjugated substrates from the UPS to macroautophagy and promotes their lysosomal degradation. Our results suggest that the Nt-Arg proteome of arginylated proteins contributes to reprogramming global proteolytic flux under stresses.
Keywords: ATE1 R-transferase; N-end rule pathway; macroautophagy; p62/STQSM/Sequestosome-1; ubiquitin-proteasome system.
Conflict of interest statement
Conflict of interest statement: Y.T.K., D.C.R., and Z.E. are coauthors on a 2016 Autophagy Guidelines publication that lists more than 1,000 authors who published their work in Autophagy.
Figures




References
-
- Deng Z, et al. Autophagy receptors and neurodegenerative diseases. Trends Cell Biol. 2017;27:491–504. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
