

Interleukins-17 and 27 promote liver regeneration by sequentially inducing progenitor cell expansion and differentiation
- PMID: 29507906
- PMCID: PMC5831061
- DOI: 10.1002/hep4.1145
Interleukins-17 and 27 promote liver regeneration by sequentially inducing progenitor cell expansion and differentiation
Abstract
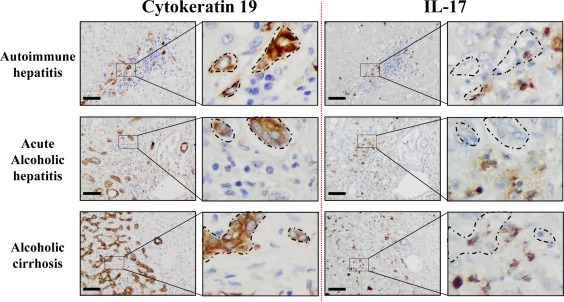
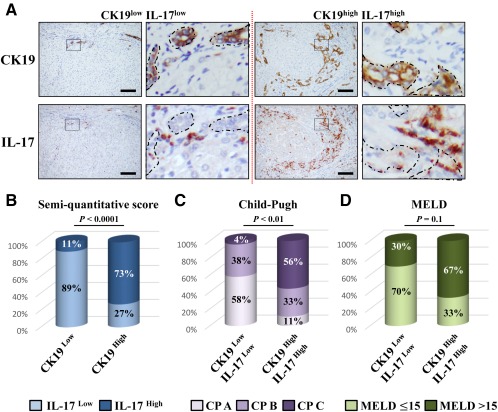
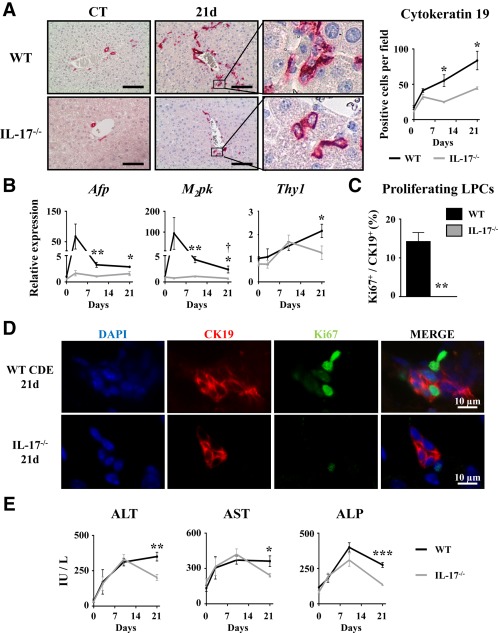
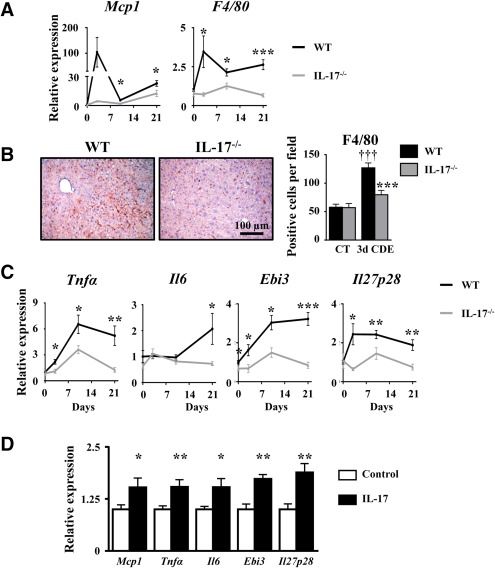
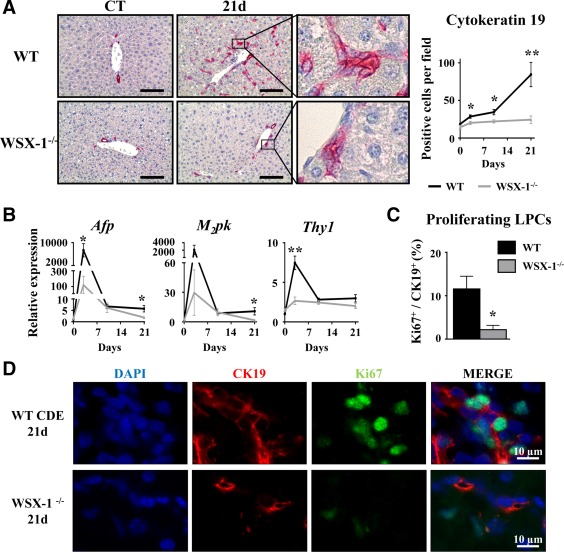
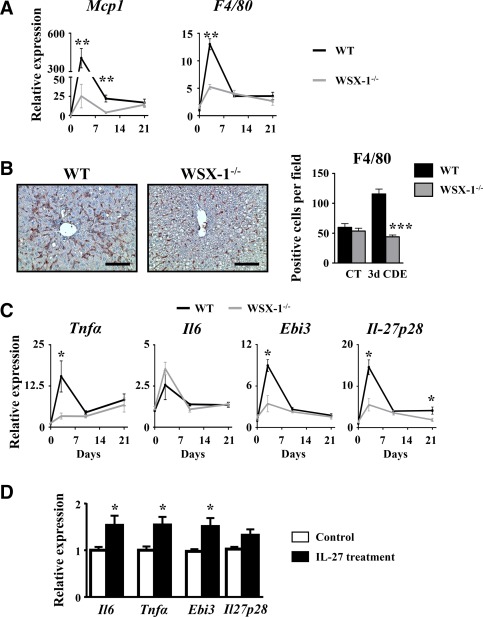
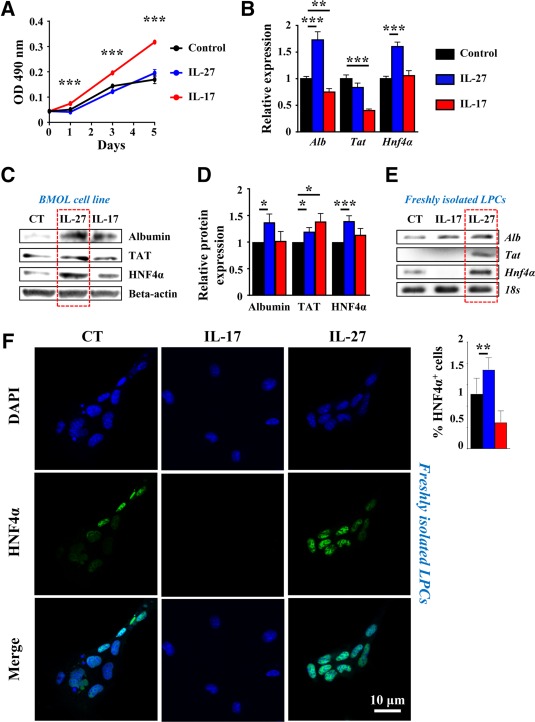
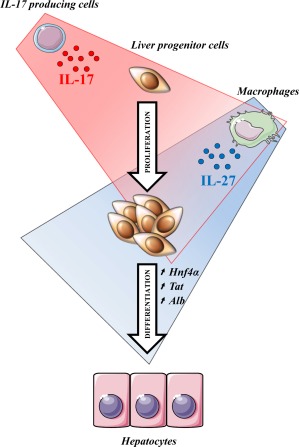
Liver progenitor cells (LPCs)/ductular reactions (DRs) are associated with inflammation and implicated in the pathogenesis of chronic liver diseases. However, how inflammation regulates LPCs/DRs remains largely unknown. Identification of inflammatory processes that involve LPC activation and expansion represent a key step in understanding the pathogenesis of liver diseases. In the current study, we found that diverse types of chronic liver diseases are associated with elevation of infiltrated interleukin (IL)-17-positive (+) cells and cytokeratin 19 (CK19)+ LPCs, and both cell types colocalized and their numbers positively correlated with each other. The role of IL-17 in the induction of LPCs was examined in a mouse model fed a choline-deficient and ethionine-supplemented (CDE) diet. Feeding of wild-type mice with the CDE diet markedly elevated CK19+Ki67+ proliferating LPCs and hepatic inflammation. Disruption of the IL-17 gene or IL-27 receptor, alpha subunit (WSX-1) gene abolished CDE diet-induced LPC expansion and inflammation. In vitro treatment with IL-17 promoted proliferation of bipotential murine oval liver cells (a liver progenitor cell line) and markedly up-regulated IL-27 expression in macrophages. Treatment with IL-27 favored the differentiation of bipotential murine oval liver cells and freshly isolated LPCs into hepatocytes. Conclusion: The current data provide evidence for a collaborative role between IL-17 and IL-27 in promoting LPC expansion and differentiation, respectively, thereby contributing to liver regeneration. (Hepatology Communications 2018;2:329-343).
Figures
References
-
- Williams MJ, Clouston AD, Forbes SJ. Links between hepatic fibrosis, ductular reaction, and progenitor cell expansion. Gastroenterology 2014;146:349‐356. - PubMed
-
- Desmet VJ. Ductal plates in hepatic ductular reactions. Hypothesis and implications. I. Types of ductular reaction reconsidered. Virchows Arch 2011;458:251‐259. - PubMed
-
- Gouw AS, Clouston AD, Theise ND. Ductular reactions in human liver: diversity at the interface. Hepatology 2011;54:1853‐1863. - PubMed
-
- Roskams TA, Libbrecht L, Desmet VJ. Progenitor cells in diseased human liver. Semin Liver Dis 2003;23:385‐396. - PubMed
-
- Clouston AD, Powell EE, Walsh MJ, Richardson MM, Demetris AJ, Jonsson JR. Fibrosis correlates with a ductular reaction in hepatitis C: roles of impaired replication, progenitor cells and steatosis. Hepatology 2005;41:809‐818. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
