

The hypoxic tumor microenvironment in vivo selects the cancer stem cell fate of breast cancer cells
- PMID: 29510720
- PMCID: PMC5840770
- DOI: 10.1186/s13058-018-0944-8
The hypoxic tumor microenvironment in vivo selects the cancer stem cell fate of breast cancer cells
Abstract
Background: Tumor hypoxia is an independent prognostic factor associated with poor patient survival. Emerging evidence suggests that hypoxia can potentially maintain or enhance the stem cell phenotype of both normal stem cells and cancer cells. However, it remains to be determined whether cell fate is regulated in vivo by the hypoxic tumor microenvironment (TME).
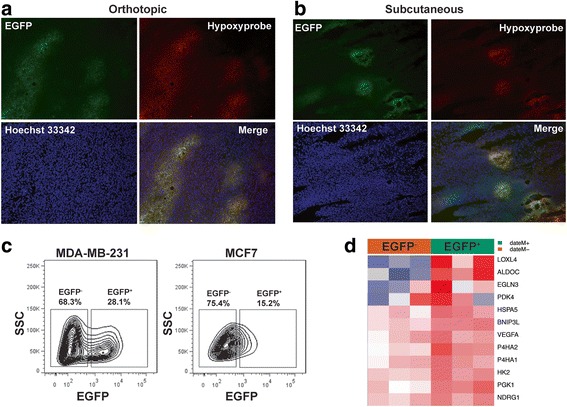
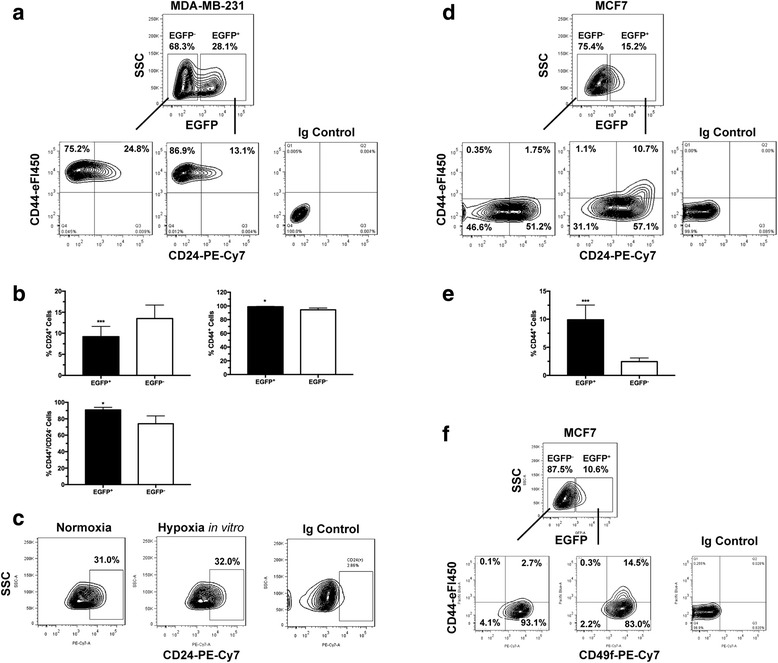
Methods: We established a hypoxia-sensing xenograft model to identify hypoxic tumor cell in vivo primarily using human breast cancer cell lines MDA-MB-231 and MCF7. Hypoxic tumor cells were identified in situ by fluorescence of green fluorescence protein. They were further isolated from xenografts, purified and sorted by flow cytometry for detailed analysis of their stem cell characteristics.
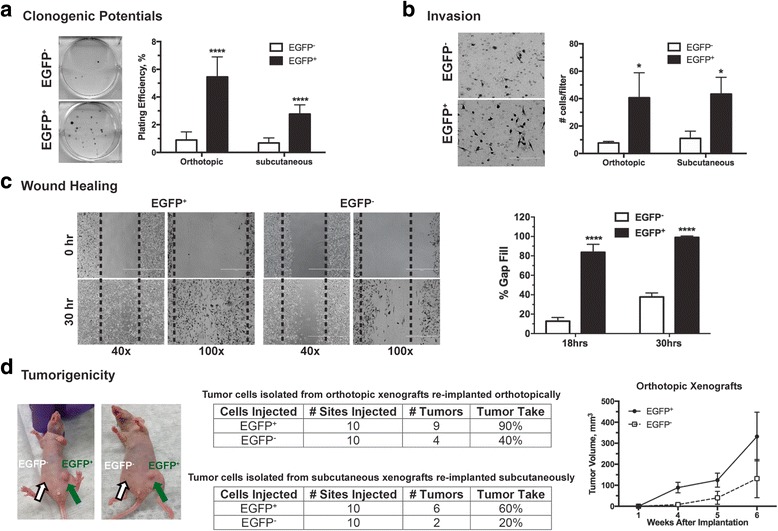
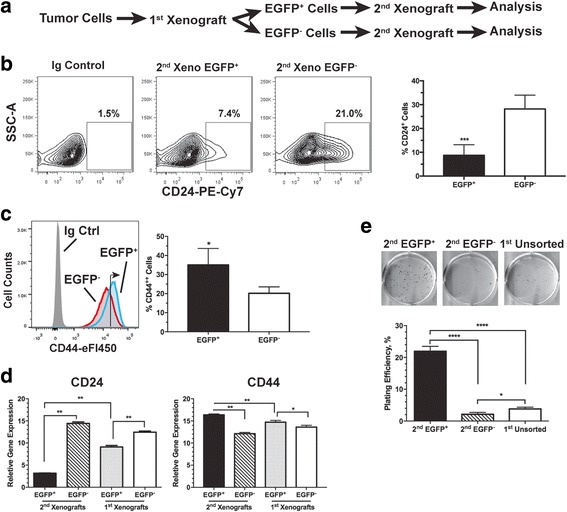
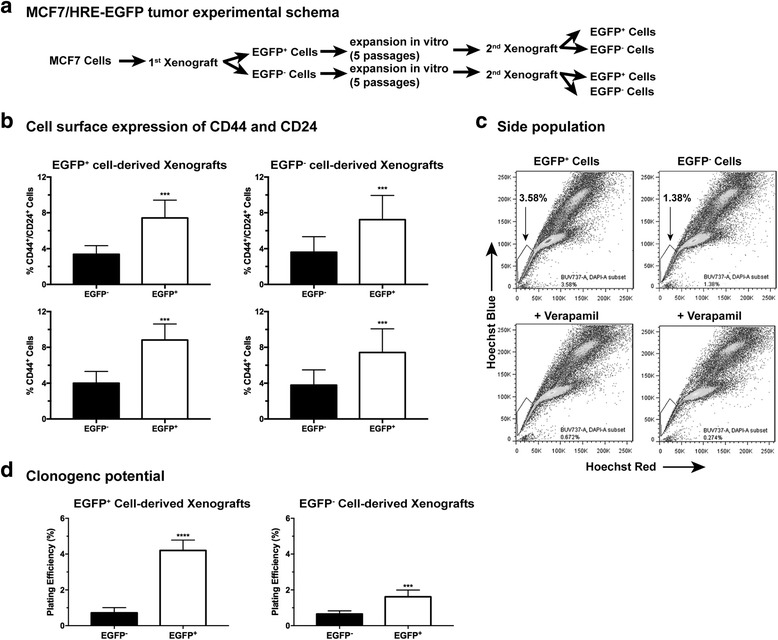
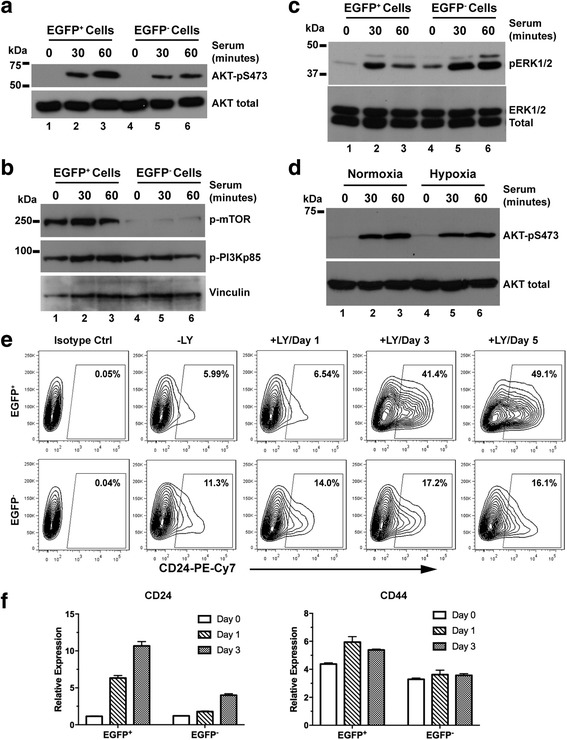
Results: We have found that hypoxic tumor cells freshly isolated from xenografts contain increased subpopulations of tumor cells with cancer stem cell (CSC)-like characteristics. The CSC characteristics of the hypoxic tumor cells are further enhanced upon re-implantation in vivo, whereas secondary xenografts derived from the non-hypoxic tumor cells remain similar to the primary xenografts. Interestingly, the phenotypes exhibited by the hypoxic tumor cells are stable and remain distinctively different from those of the non-hypoxic tumor cells isolated from the same tumor mass even when they are maintained under the same ambient culture conditions. Mechanistically, the PI3K/AKT pathway is strongly potentiated in the hypoxic tumor cells and is required to maintain the CSC-like phenotype. Importantly, the differential cell fates between hypoxic and non-hypoxic tumor cells are only found in tumor cells isolated from the hypoxic TME in vivo and are not seen in tumor cells treated by hypoxia in vitro alone.
Conclusions: These previously unknown observations suggest that the hypoxic TME may promote malignant progression and therapy resistance by coordinating induction, selection and/or preferential maintenance of the CSC-like phenotype in tumor cells.
Keywords: AKT; Breast cancer cell; Cancer stem cell; Cell fate; Hypoxia; PI3K; Tumor microenvironment; Xenograft.
Conflict of interest statement
Ethics approval and consent to participate
All animal procedures were reviewed and approved by the Institutional Animal Care & Use Committee (IACUC) of Yale University.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- Hockel M, Schlenger K, Aral B, Mitze M, Schaffer U, Vaupel P. Association between tumor hypoxia and malignant progression in advanced cancer of the uterine cervix. Cancer Res. 1996;56(19):4509–4515. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
