

Unveiling skin macrophage dynamics explains both tattoo persistence and strenuous removal
- PMID: 29511065
- PMCID: PMC5881467
- DOI: 10.1084/jem.20171608
Unveiling skin macrophage dynamics explains both tattoo persistence and strenuous removal
Abstract
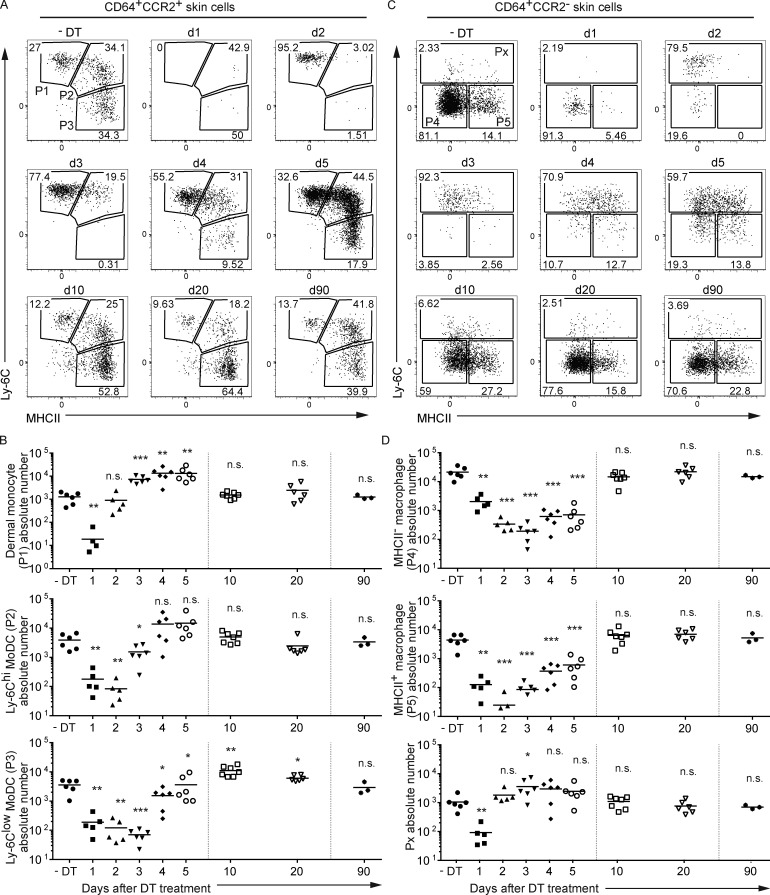
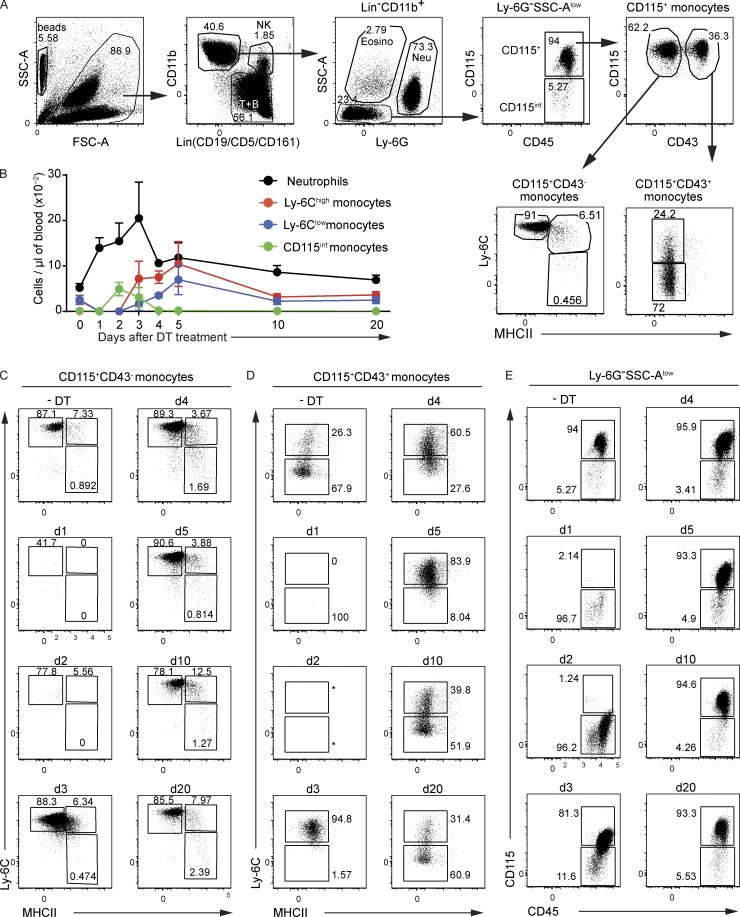
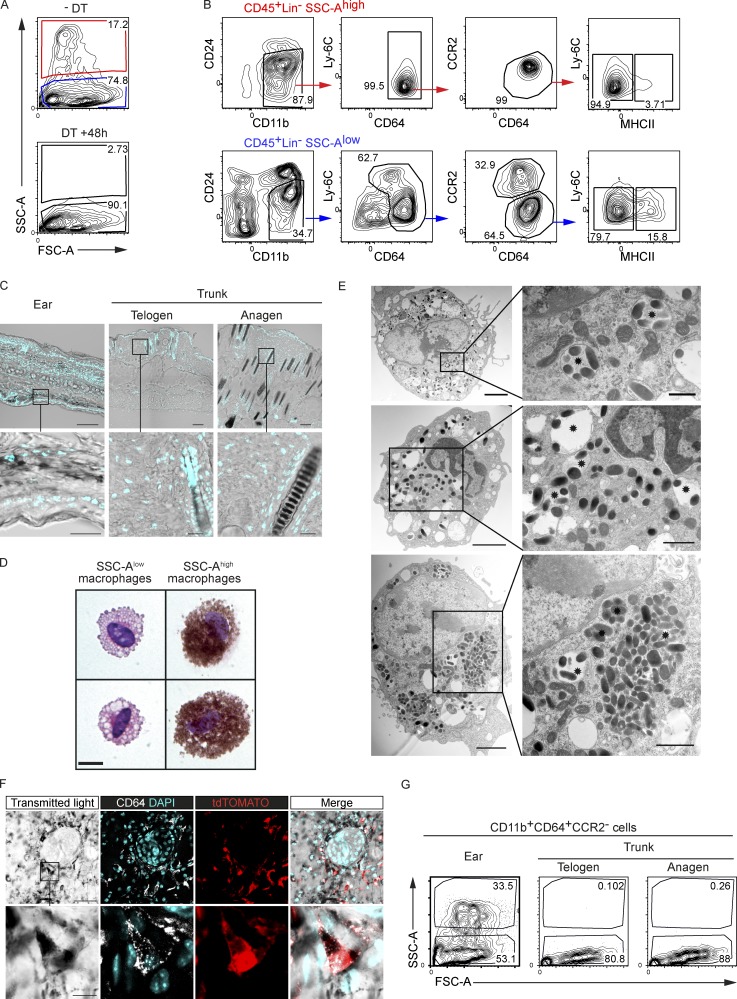
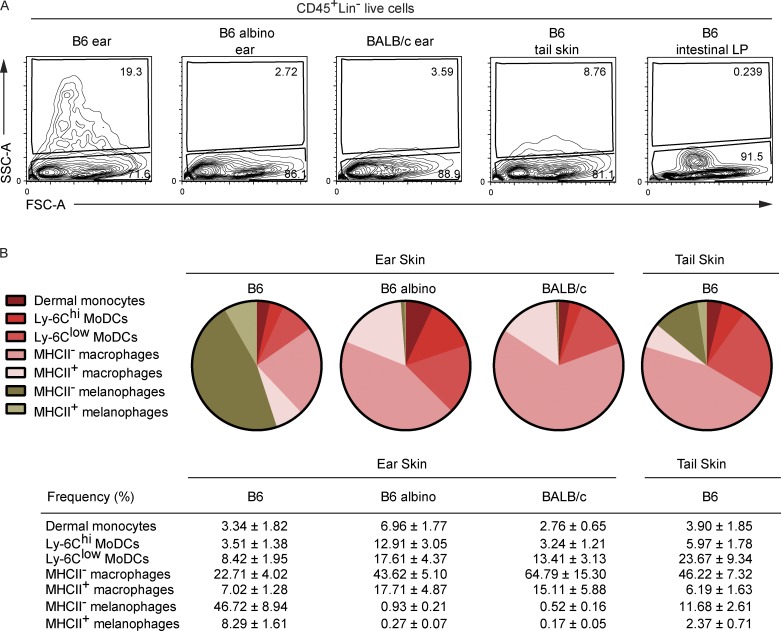
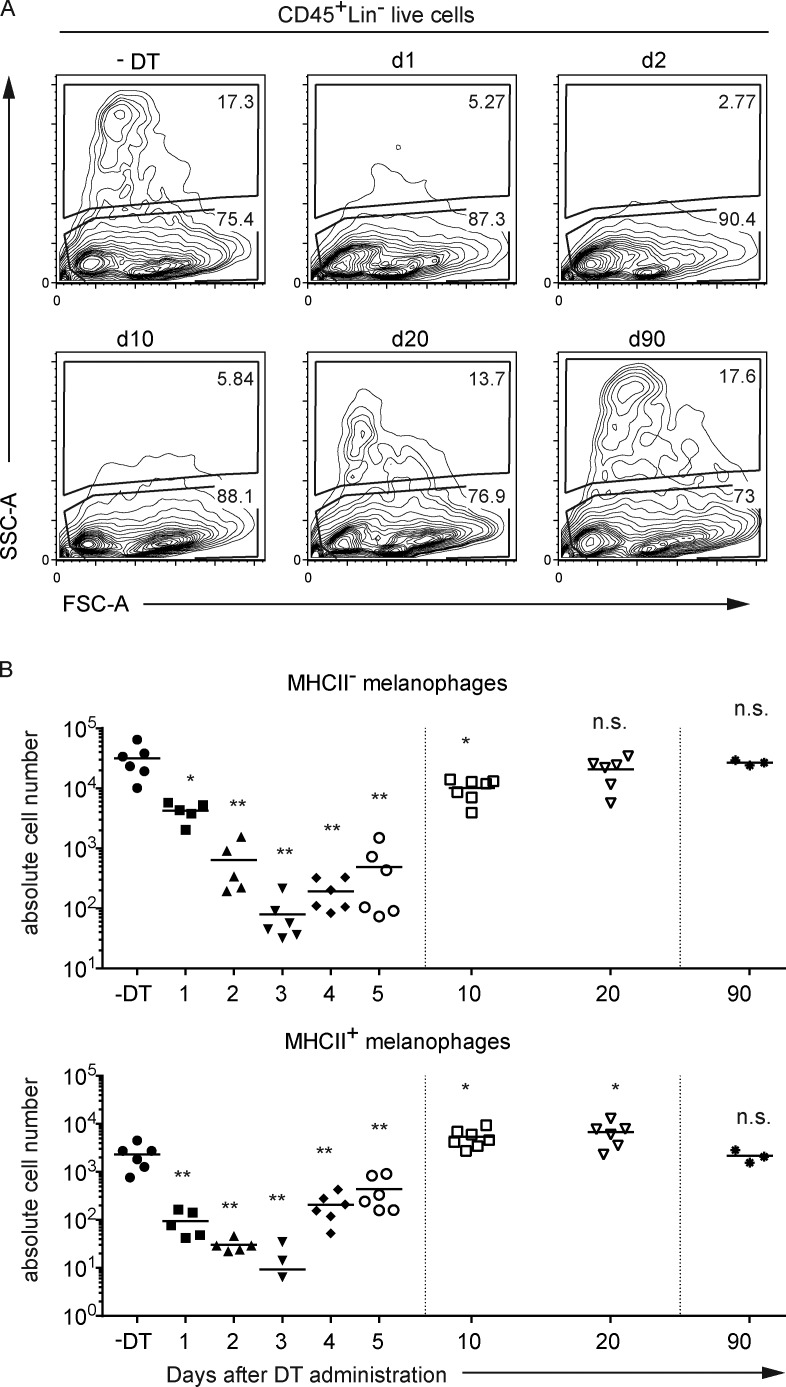
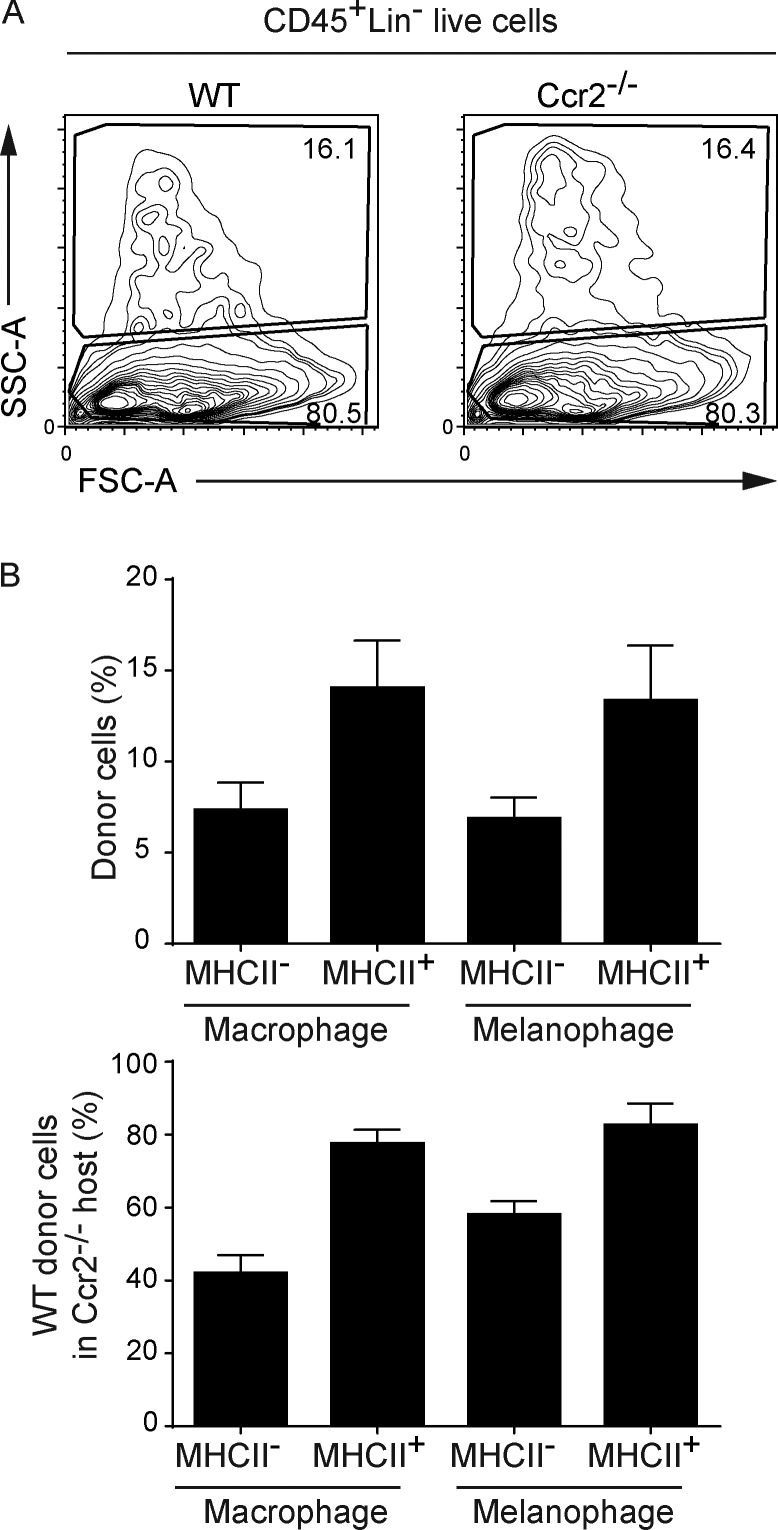
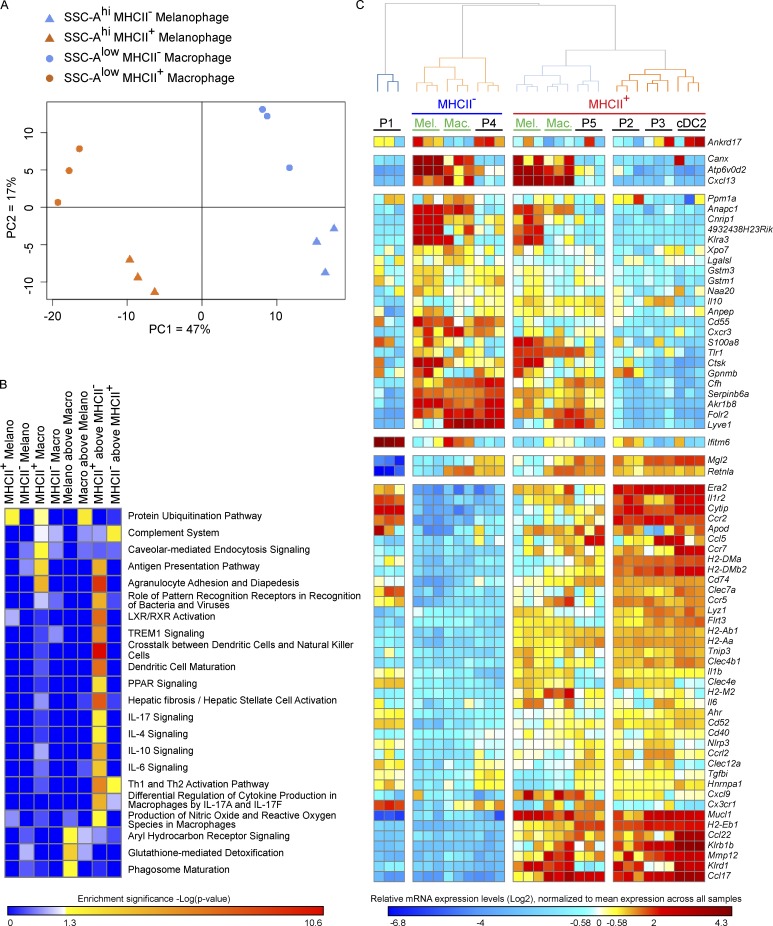
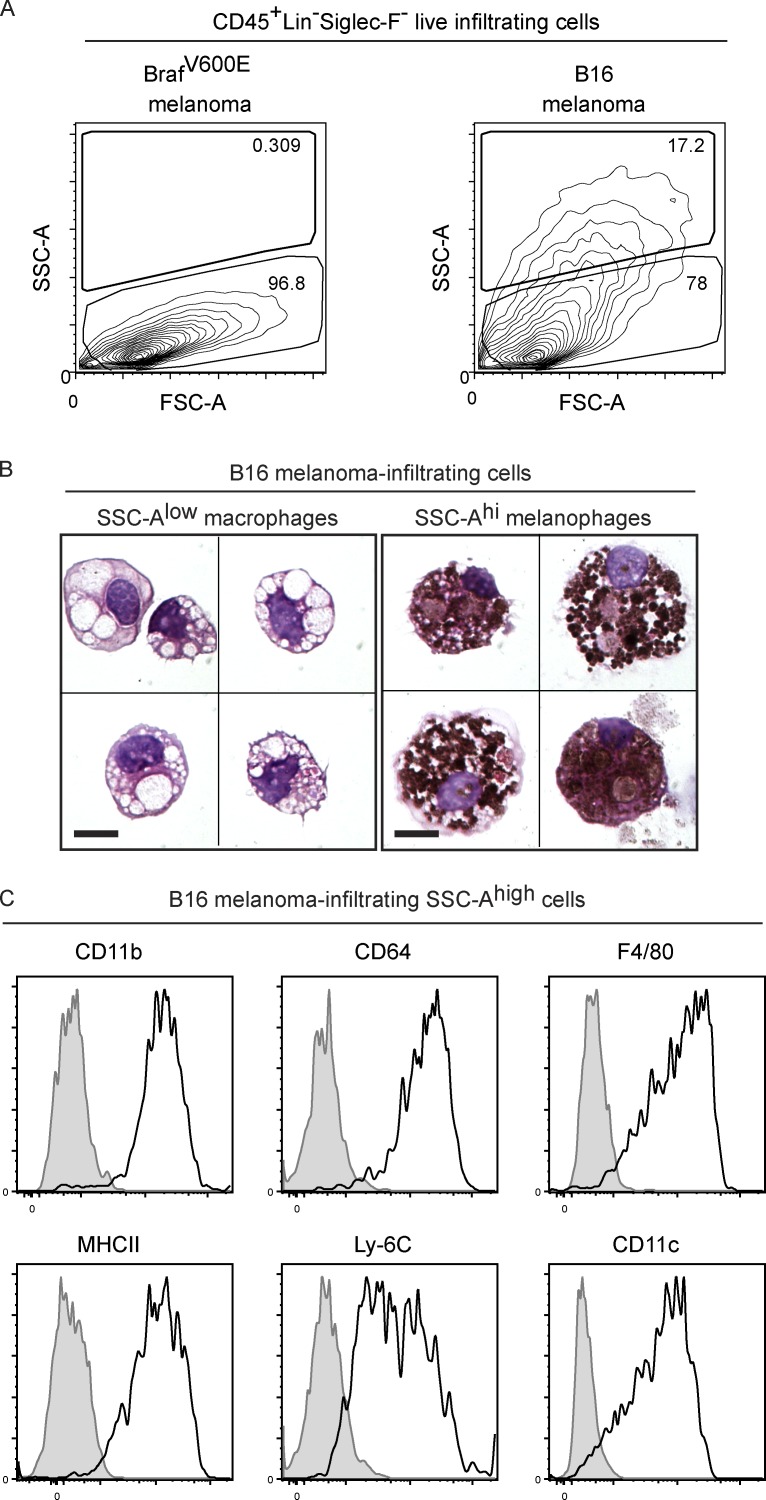
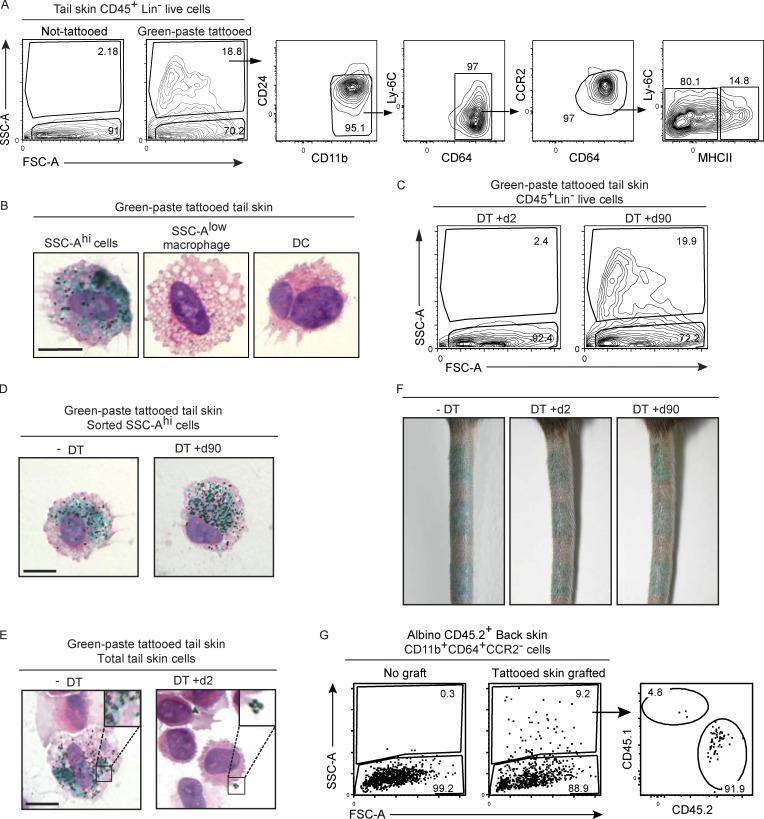
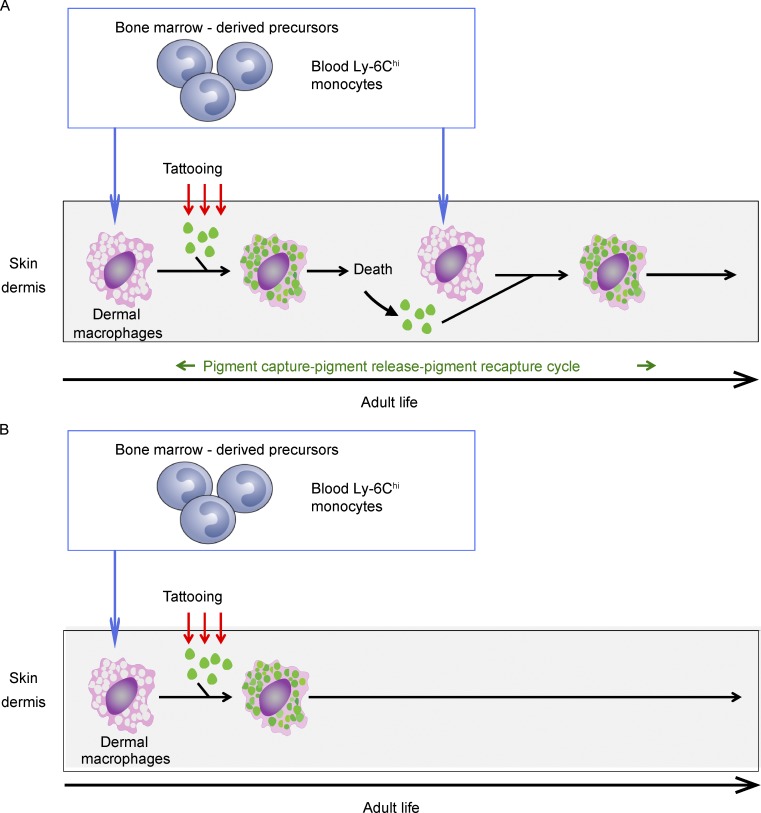
Here we describe a new mouse model that exploits the pattern of expression of the high-affinity IgG receptor (CD64) and allows diphtheria toxin (DT)-mediated ablation of tissue-resident macrophages and monocyte-derived cells. We found that the myeloid cells of the ear skin dermis are dominated by DT-sensitive, melanin-laden cells that have been missed in previous studies and correspond to macrophages that have ingested melanosomes from neighboring melanocytes. Those cells have been referred to as melanophages in humans. We also identified melanophages in melanocytic melanoma. Benefiting of our knowledge on melanophage dynamics, we determined the identity, origin, and dynamics of the skin myeloid cells that capture and retain tattoo pigment particles. We showed that they are exclusively made of dermal macrophages. Using the possibility to delete them, we further demonstrated that tattoo pigment particles can undergo successive cycles of capture-release-recapture without any tattoo vanishing. Therefore, congruent with dermal macrophage dynamics, long-term tattoo persistence likely relies on macrophage renewal rather than on macrophage longevity.
© 2018 Baranska et al.
Figures
Comment in
-
Death, eaters, and dark marks.J Exp Med. 2018 Apr 2;215(4):1005-1006. doi: 10.1084/jem.20180311. Epub 2018 Mar 20. J Exp Med. 2018. PMID: 29559503 Free PMC article.
References
-
- Autenrieth S.E., Warnke P., Wabnitz G.H., Lucero Estrada C., Pasquevich K.A., Drechsler D., Günter M., Hochweller K., Novakovic A., Beer-Hammer S., et al. 2012. Depletion of dendritic cells enhances innate anti-bacterial host defense through modulation of phagocyte homeostasis. PLoS Pathog. 8:e1002552 10.1371/journal.ppat.1002552 - DOI - PMC - PubMed
-
- Bain C.C., Scott C.L., Uronen-Hansson H., Gudjonsson S., Jansson O., Grip O., Guilliams M., Malissen B., Agace W.W., and Mowat A.M.. 2013. Resident and pro-inflammatory macrophages in the colon represent alternative context-dependent fates of the same Ly6Chi monocyte precursors. Mucosal Immunol. 6:498–510. 10.1038/mi.2012.89 - DOI - PMC - PubMed
-
- Bain C.C., Bravo-Blas A., Scott C.L., Perdiguero E.G., Geissmann F., Henri S., Malissen B., Osborne L.C., Artis D., and Mowat A.M.. 2014. Constant replenishment from circulating monocytes maintains the macrophage pool in the intestine of adult mice. Nat. Immunol. 15:929–937. 10.1038/ni.2967 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
