

Herbivorous turtle ants obtain essential nutrients from a conserved nitrogen-recycling gut microbiome
- PMID: 29511180
- PMCID: PMC5840417
- DOI: 10.1038/s41467-018-03357-y
Herbivorous turtle ants obtain essential nutrients from a conserved nitrogen-recycling gut microbiome
Erratum in
-
Author Correction: Herbivorous turtle ants obtain essential nutrients from a conserved nitrogen-recycling gut microbiome.Nat Commun. 2018 Jun 19;9(1):2440. doi: 10.1038/s41467-018-04935-w. Nat Commun. 2018. PMID: 29921959 Free PMC article.
Abstract
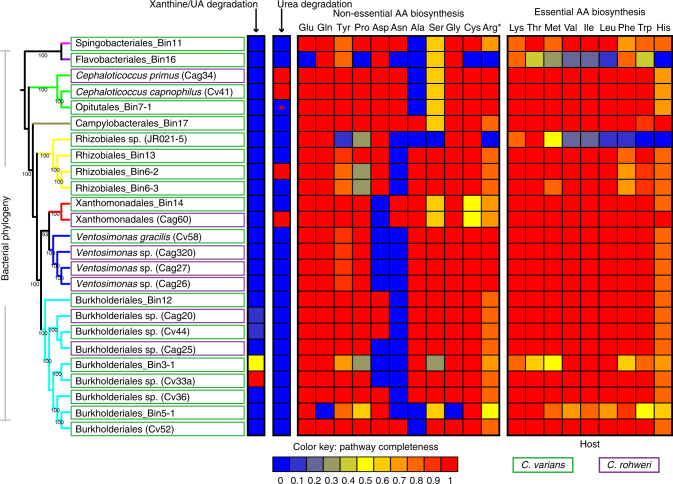
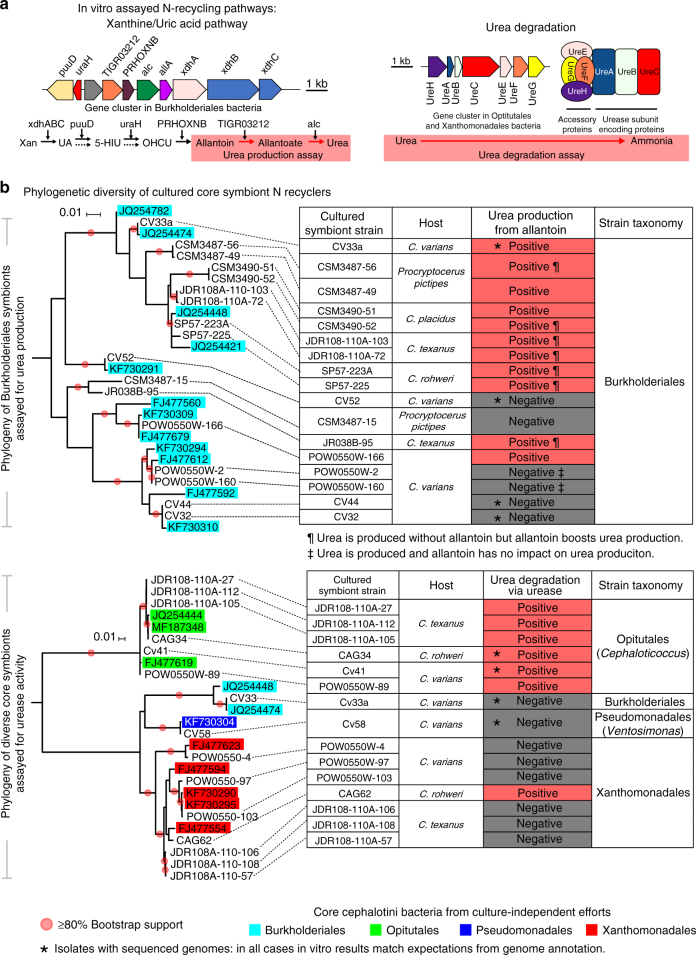
Nitrogen acquisition is a major challenge for herbivorous animals, and the repeated origins of herbivory across the ants have raised expectations that nutritional symbionts have shaped their diversification. Direct evidence for N provisioning by internally housed symbionts is rare in animals; among the ants, it has been documented for just one lineage. In this study we dissect functional contributions by bacteria from a conserved, multi-partite gut symbiosis in herbivorous Cephalotes ants through in vivo experiments, metagenomics, and in vitro assays. Gut bacteria recycle urea, and likely uric acid, using recycled N to synthesize essential amino acids that are acquired by hosts in substantial quantities. Specialized core symbionts of 17 studied Cephalotes species encode the pathways directing these activities, and several recycle N in vitro. These findings point to a highly efficient N economy, and a nutritional mutualism preserved for millions of years through the derived behaviors and gut anatomy of Cephalotes ants.
Conflict of interest statement
The authors declare no competing interests.
Figures
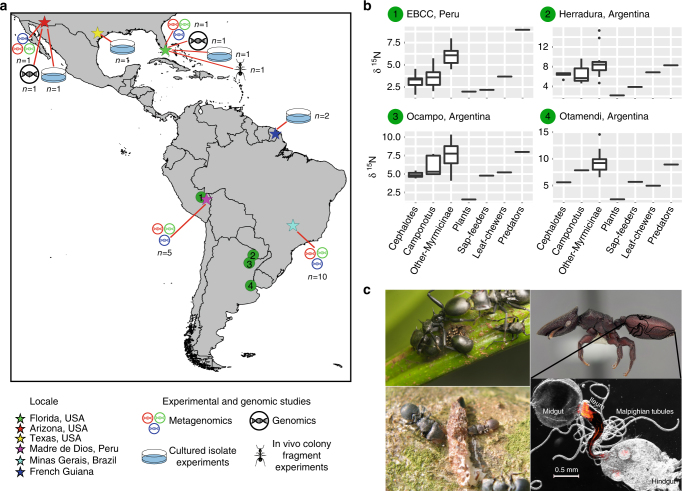
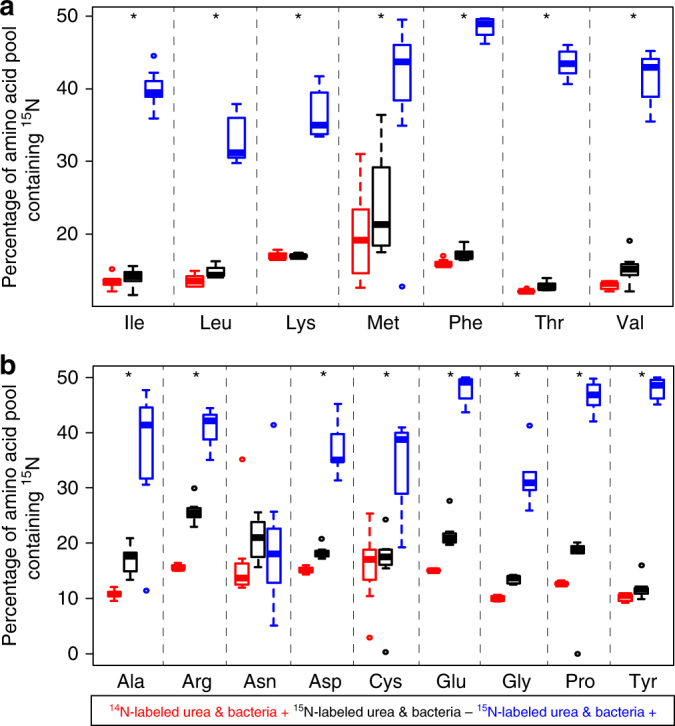
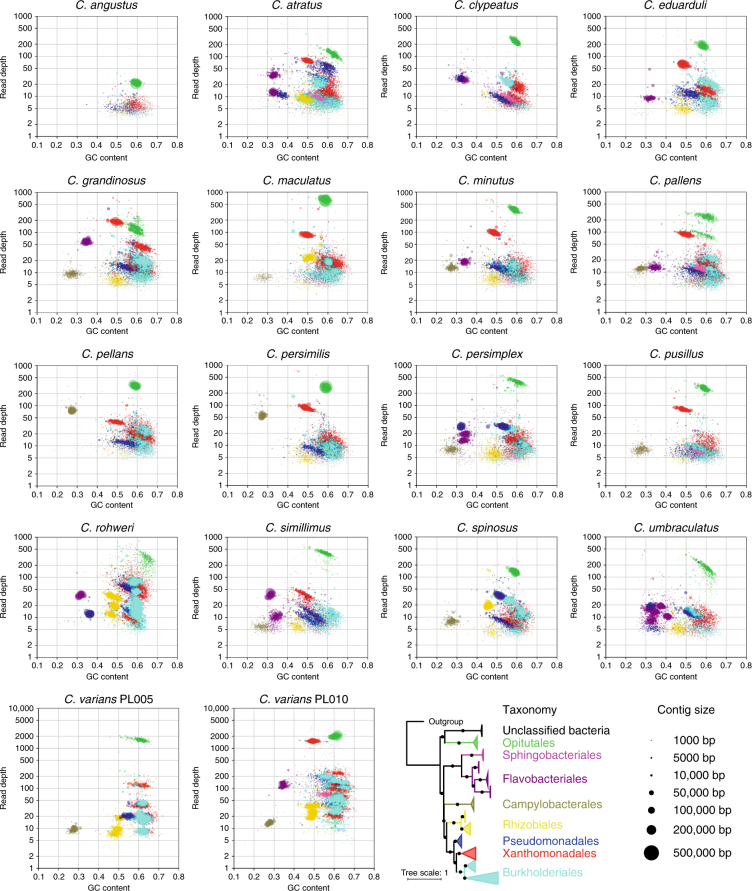
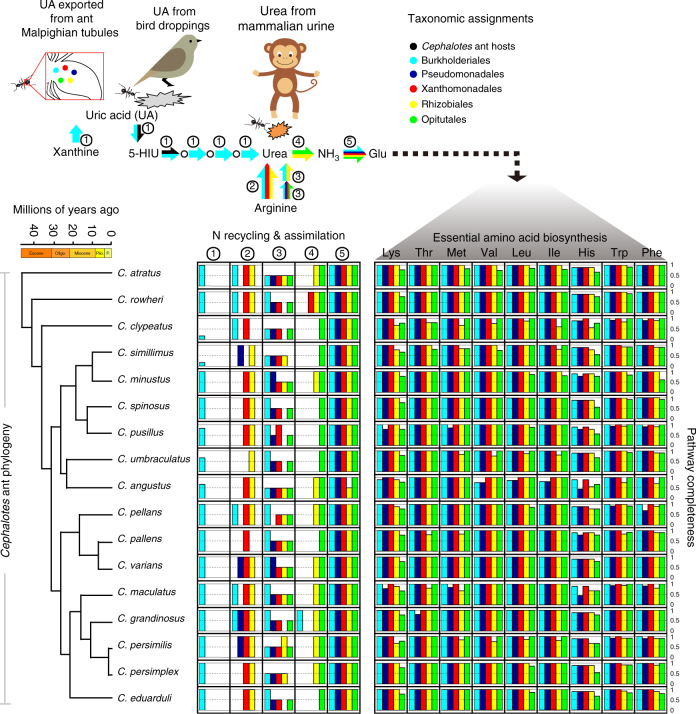
References
-
- Chapin FS. The mineral nutrition of wild plants. Annu. Rev. Ecol. Syst. 1980;11:233–260. doi: 10.1146/annurev.es.11.110180.001313. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
