

Post-translational modification of OCT4 in breast cancer tumorigenesis
- PMID: 29511337
- PMCID: PMC6180041
- DOI: 10.1038/s41418-018-0079-6
Post-translational modification of OCT4 in breast cancer tumorigenesis
Erratum in
-
Correction to: Post-translational modification of OCT4 in breast cancer tumorigenesis.Cell Death Differ. 2020 May;27(5):1743. doi: 10.1038/s41418-019-0439-x. Cell Death Differ. 2020. PMID: 31641242 Free PMC article.
Abstract
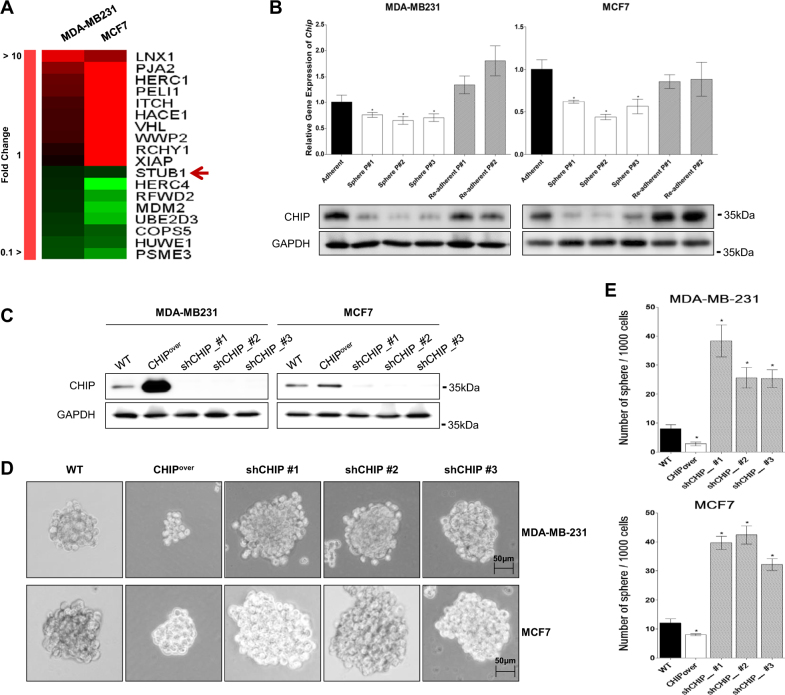
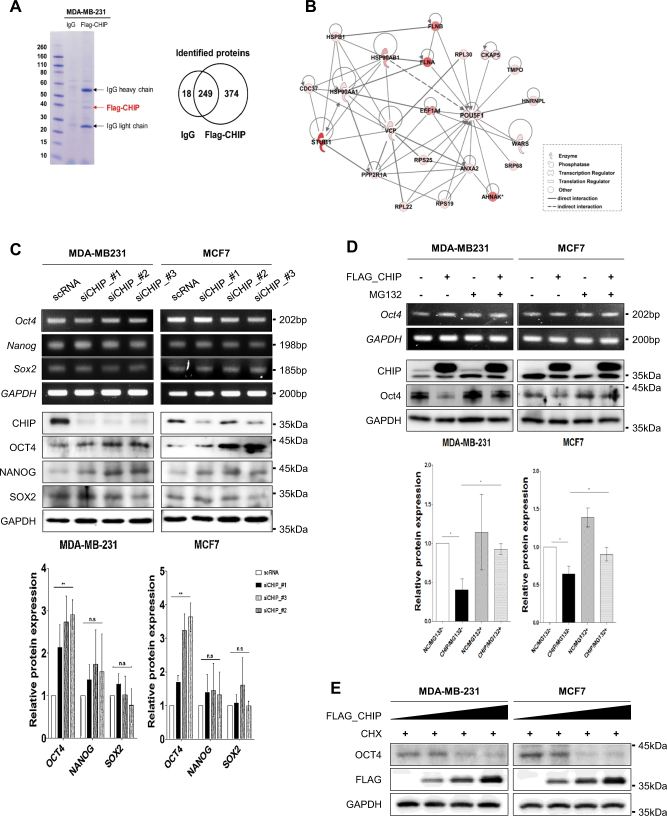
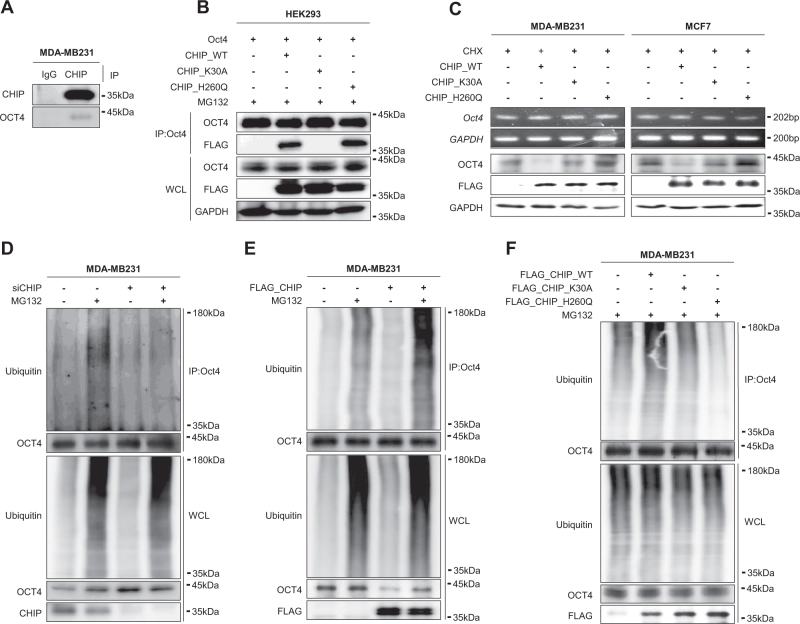
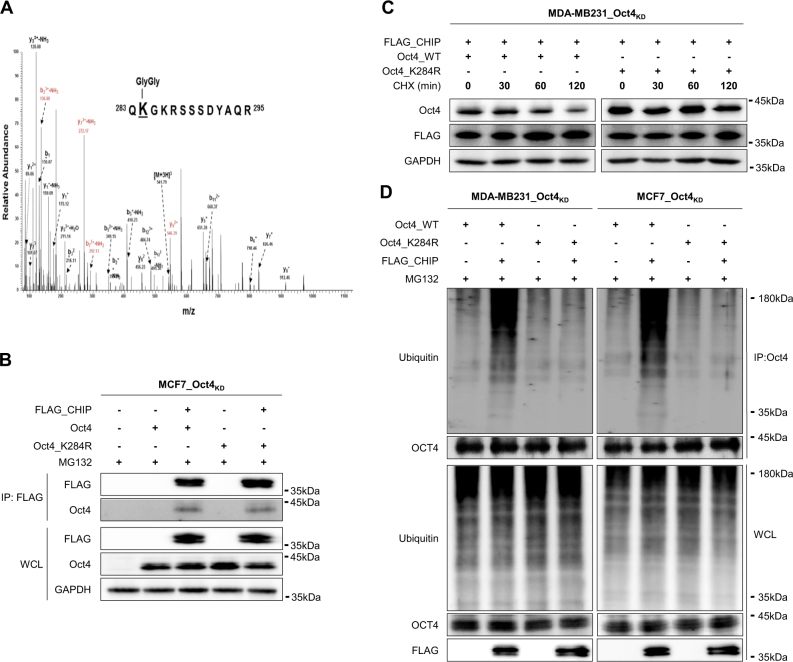
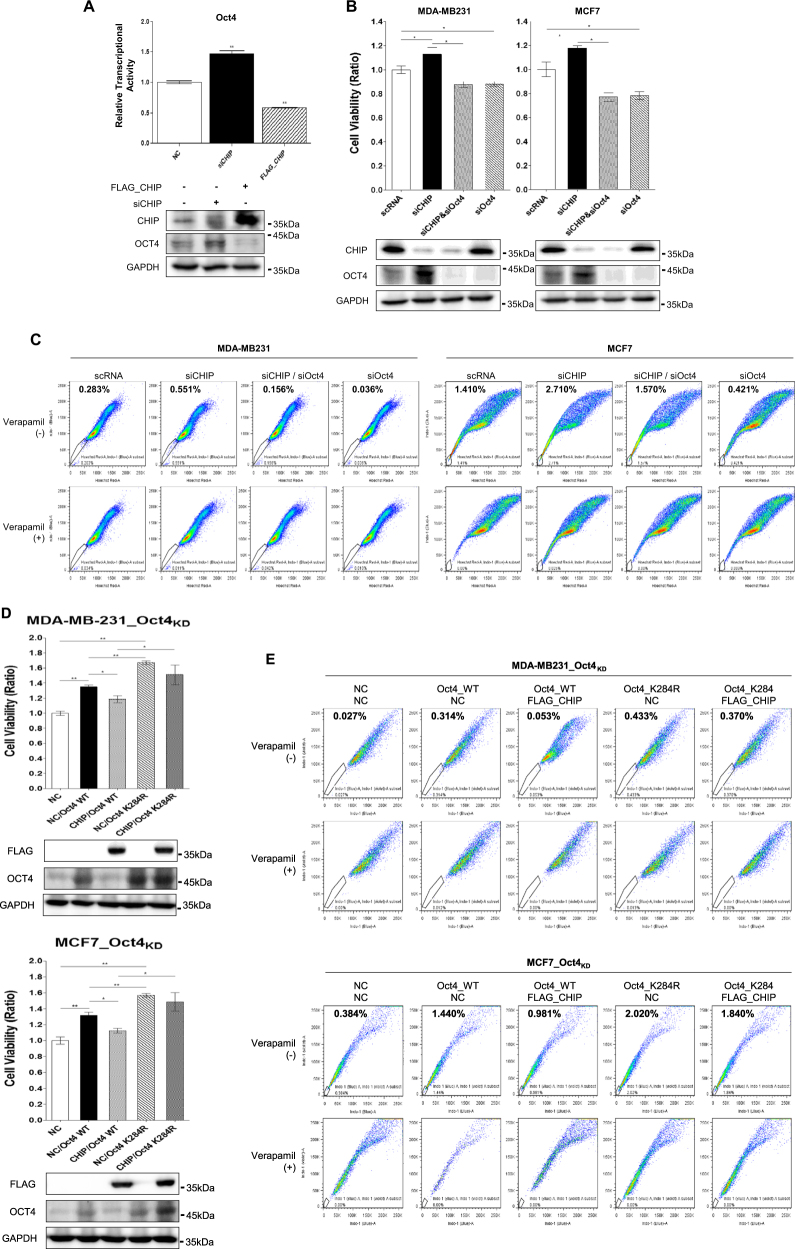
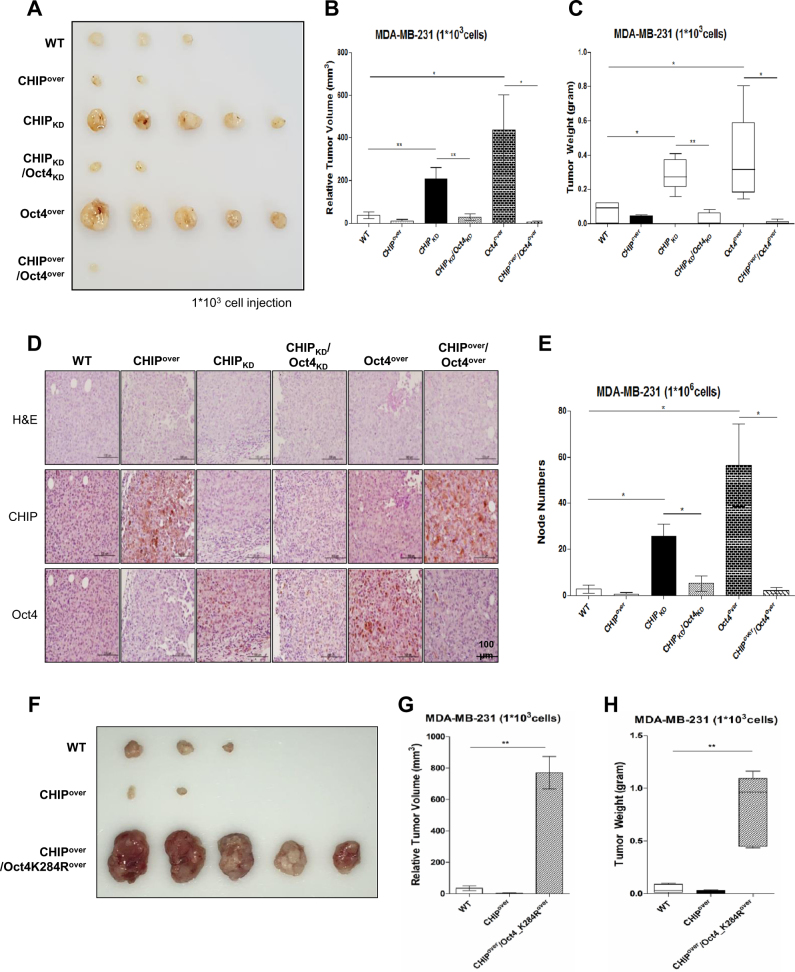
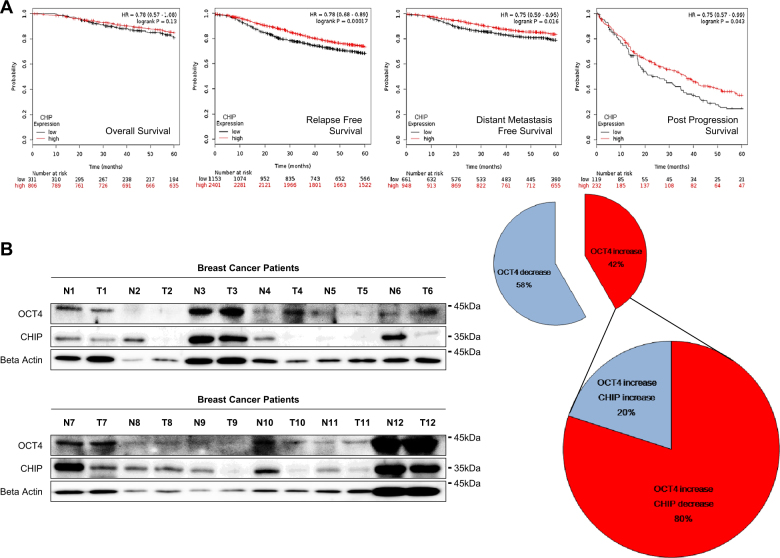
Recurrence and drug resistance of breast cancer are still the main reasons for breast cancer-associated deaths. Cancer stem cell (CSC) model has been proposed as a hypothesis for the lethality of breast cancer. Molecular mechanisms underlying CSC maintenance are still unclear. In this study, we generated mammospheres derived from breast cancer MDA-MB231 cells and MCF7 cells to enrich CSCs and performed DNA microarray analysis. We found that the expression of carboxy terminus of HSP70-interacting protein (CHIP) E3 ubiquitin ligase was significantly downregulated in breast CSCs. CHIP depletion increased mammosphere formation, whereas CHIP overexpression reversed this effect. We identified interactomes by mass spectrometry and detected CHIP directly interacted with OCT4, a stemness factor. CHIP overexpression decreased OCT4 stability through proteasomal degradation. CHIP induced OCT4 ubiquitination, whereas H260Q, a catalytic CHIP mutant, did not. Interestingly, we determined that OCT4 was ubiquitinated at lysine 284, and CHIP overexpression did not degrade K284R mutant OCT4. CHIP overexpression decreased the proliferation and side population of breast cancer cells, but these were not occurred in K284R mutant OCT4 overexpressed cells. Only 1000 cells showing CHIP depletion or OCT4 overexpression sufficiently generated breast tumors and lung metastases in xenografted mice. Ubiquitination-defective mutant of OCT4(K284R) overexpressed cells drastically generated tumor burdens in mice. Patients with breast cancer who showed low CHIP expression had poor survival probability. Taken together, we suggest that CHIP-induced OCT4 ubiquitination is important in breast CSCs. Regulation of CHIP expression and OCT4 protein stability is a considerable approach for breast cancer therapy.
Conflict of interest statement
:The authors declare that they have no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
