

Paracrine Interactions within the Pancreatic Islet Determine the Glycemic Set Point
- PMID: 29514065
- PMCID: PMC5872154
- DOI: 10.1016/j.cmet.2018.01.015
Paracrine Interactions within the Pancreatic Islet Determine the Glycemic Set Point
Abstract
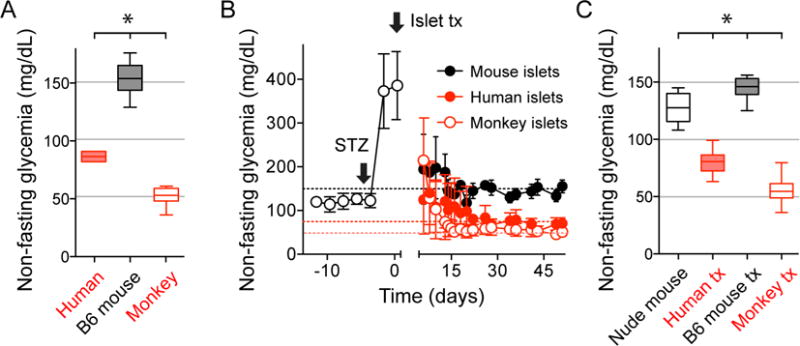
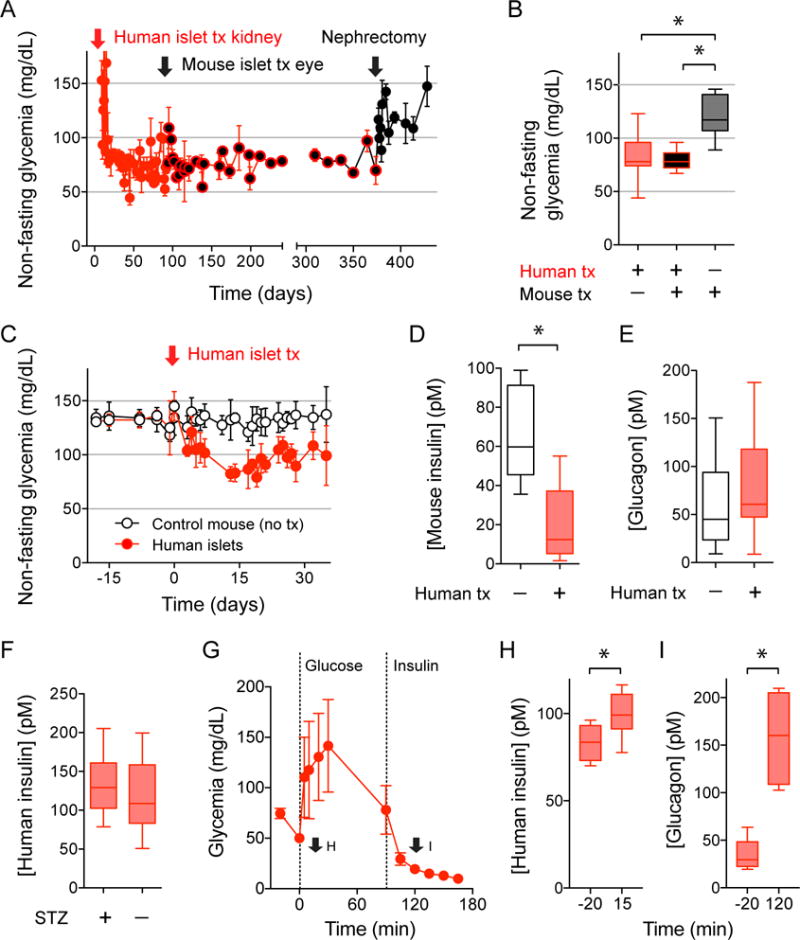
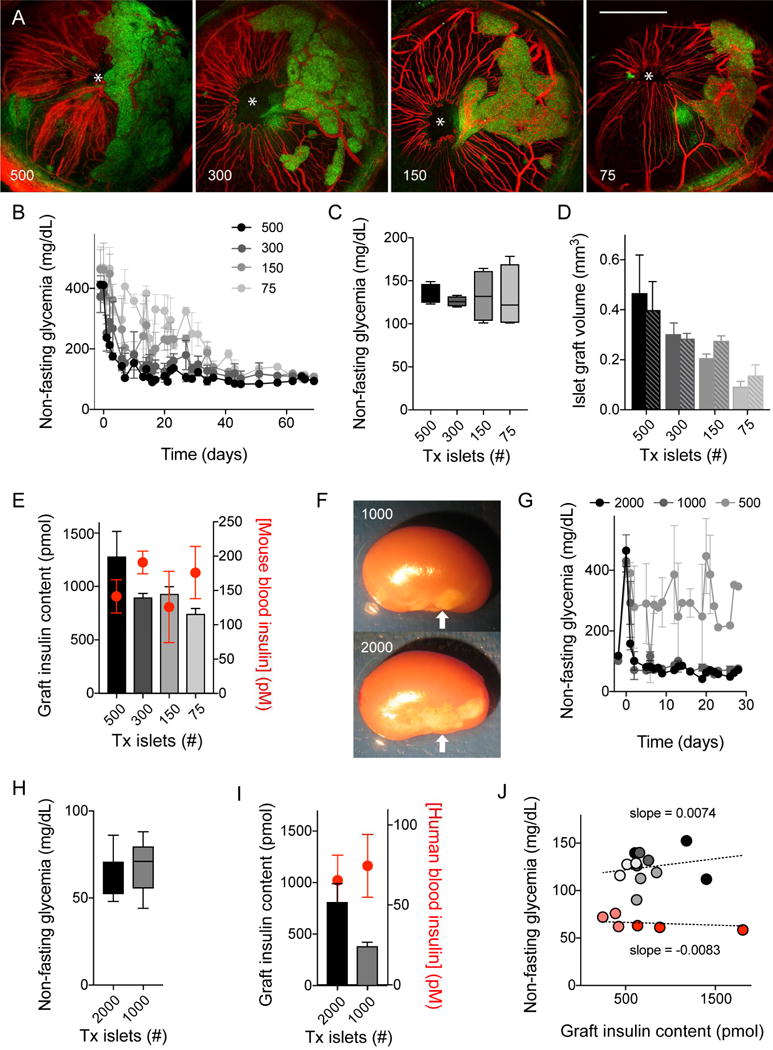
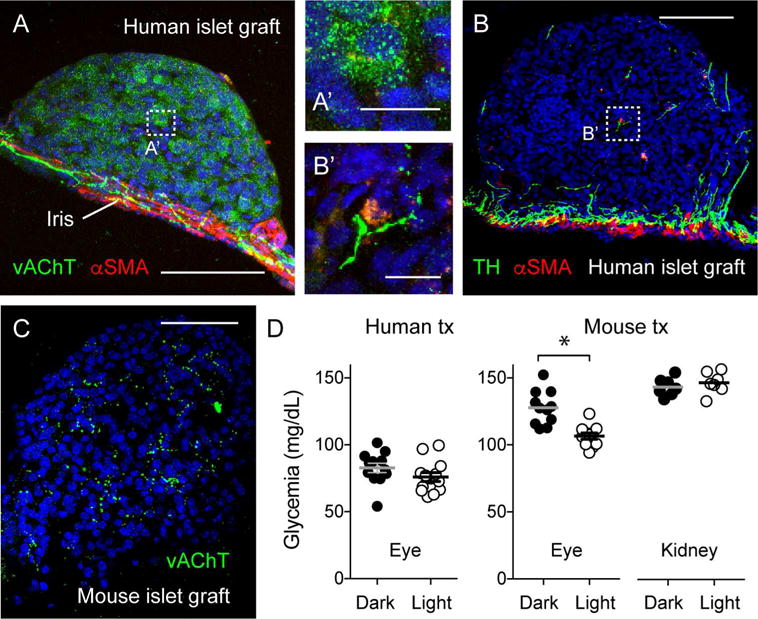
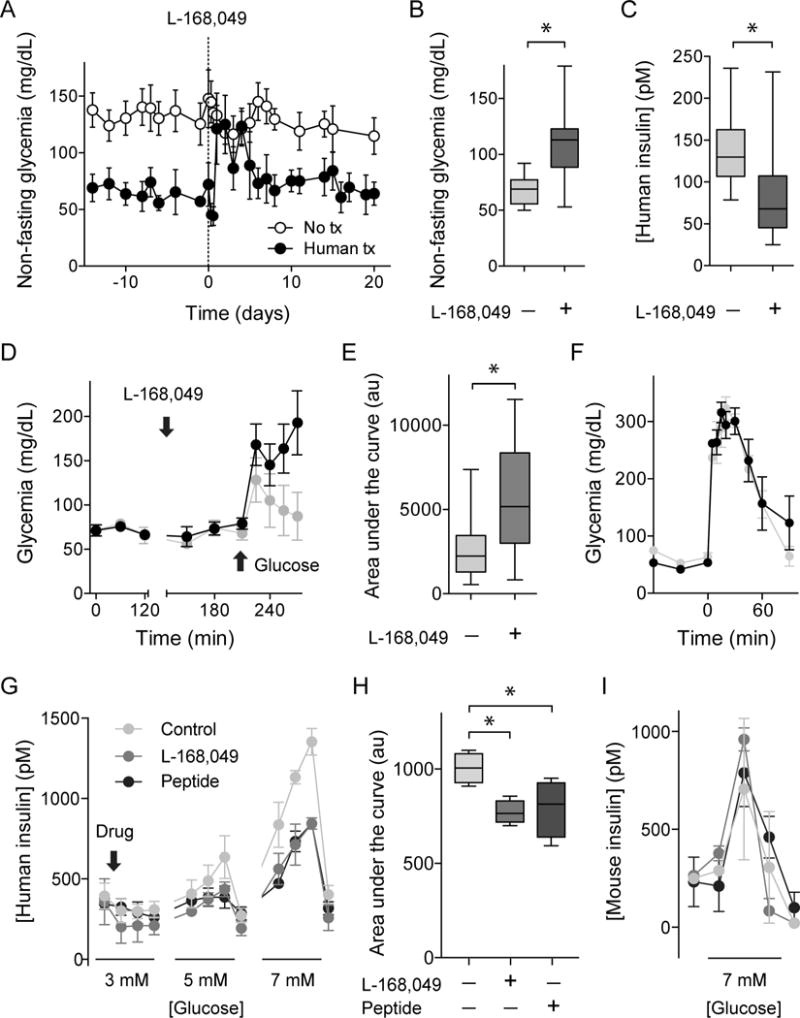
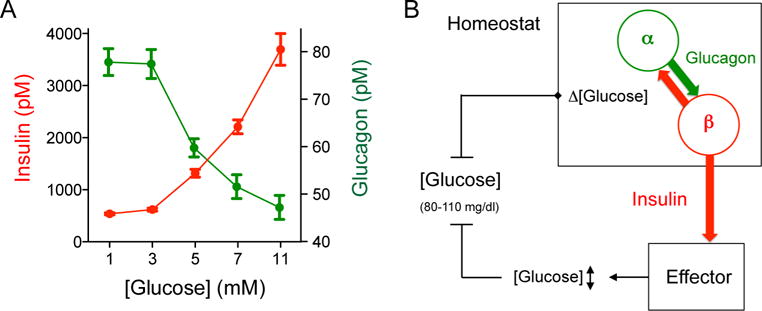
Every animal species has a signature blood glucose level or glycemic set point. These set points are different, and the normal glycemic levels (normoglycemia) of one species would be life threatening for other species. Mouse normoglycemia can be considered diabetic for humans. The biological determinants of the glycemic set point remain unclear. Here we show that the pancreatic islet imposes its glycemic set point on the organism, making it the bona fide glucostat in the body. Moreover, and in contrast to rodent islets, glucagon input from the alpha cell to the insulin-secreting beta cell is necessary to fine-tune the distinctive human set point. These findings affect transplantation and regenerative approaches to treat diabetes because restoring normoglycemia may require more than replacing only the beta cells. Furthermore, therapeutic strategies using glucagon receptor antagonists as hypoglycemic agents need to be reassessed, as they may reset the overall glucostat in the organism.
Keywords: alpha cell; beta cell; glucagon; glucose homeostasis; glycemia; glycemic set point; human pancreatic islet; humanized mouse; insulin secretion; paracrine signaling.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
P.-O.B. is cofounder and CEO of Biocrine, an unlisted biotech company that is using the anterior chamber of the eye technique as a research tool. B.L., I.L., and M.H.A. are consultants for the same company.
A.P. is currently employed at the National Institutes of Health (NIH). The opinions expressed in this article are the author’s own and do not necessarily reflect the views of the National Institutes of Health, the Department of Health and Human Services, or the United States government.
Figures
References
-
- Bertuzzi F, Berra C, Socci C, Davalli AM, Calori G, Freschi M, Piemonti L, De Nittis P, Pozza G, Pontiroli AE. Glucagon improves insulin secretion from pig islets in vitro. J Endocrinol. 1995;147:87–93. - PubMed
-
- Bottino R, Inverardi L, Valente U, Ricordi C. Serum-free medium and pyruvate improve survival and glucose responsiveness of islet beta cells in culture. Transplant Proc. 1997;29:1978. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
