

α Cell Function and Gene Expression Are Compromised in Type 1 Diabetes
- PMID: 29514095
- PMCID: PMC6368357
- DOI: 10.1016/j.celrep.2018.02.032
α Cell Function and Gene Expression Are Compromised in Type 1 Diabetes
Abstract
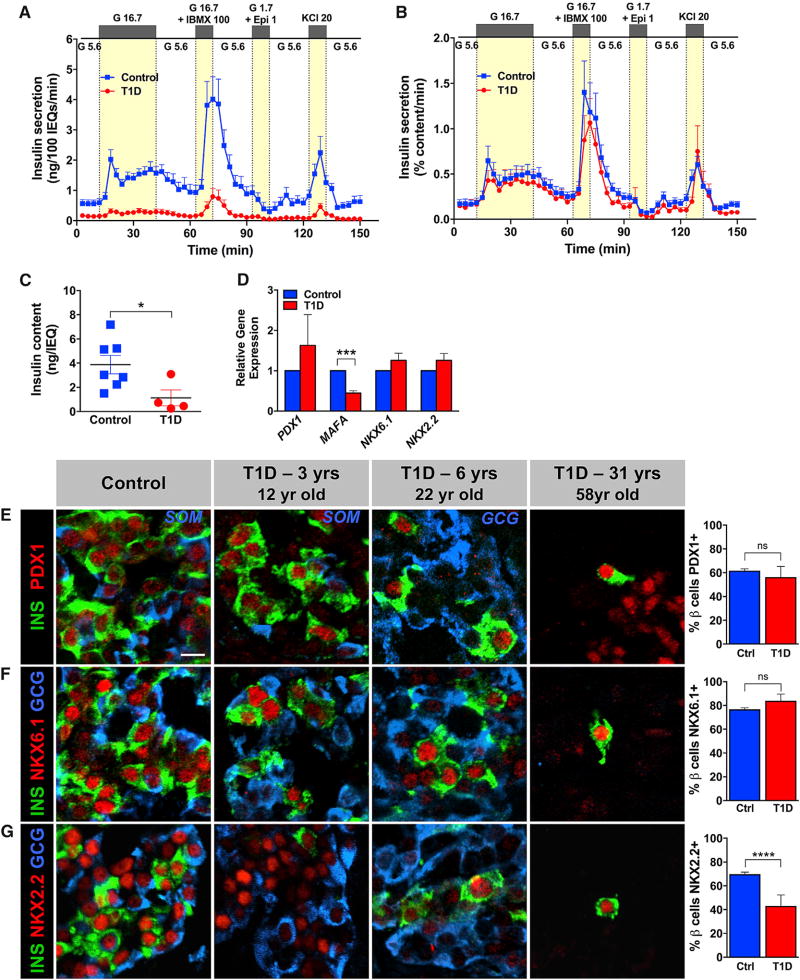
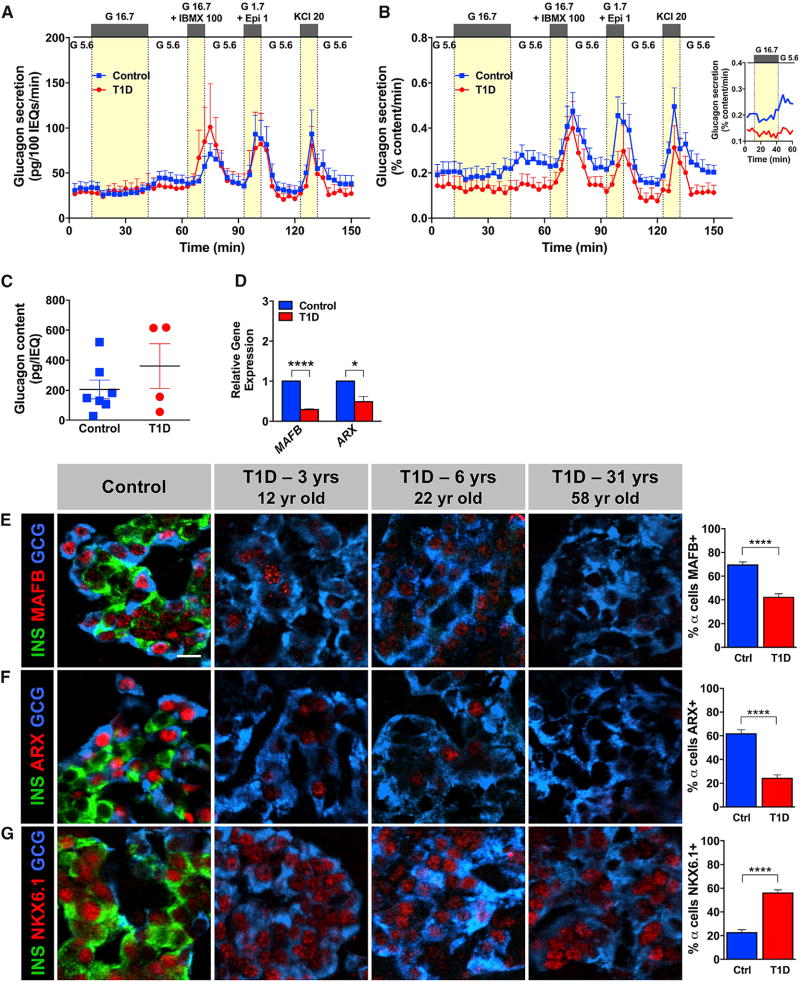
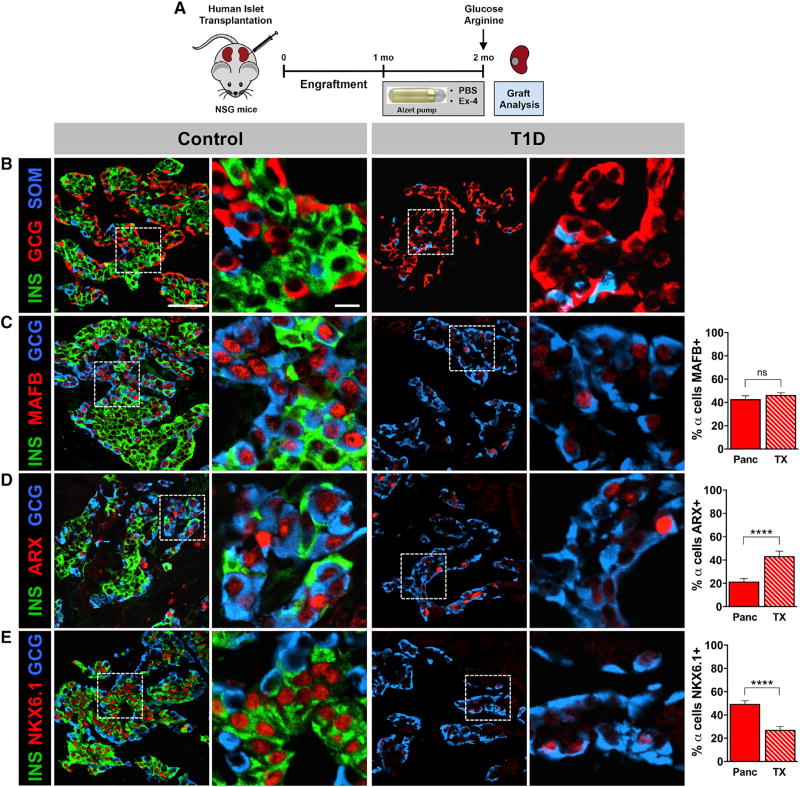
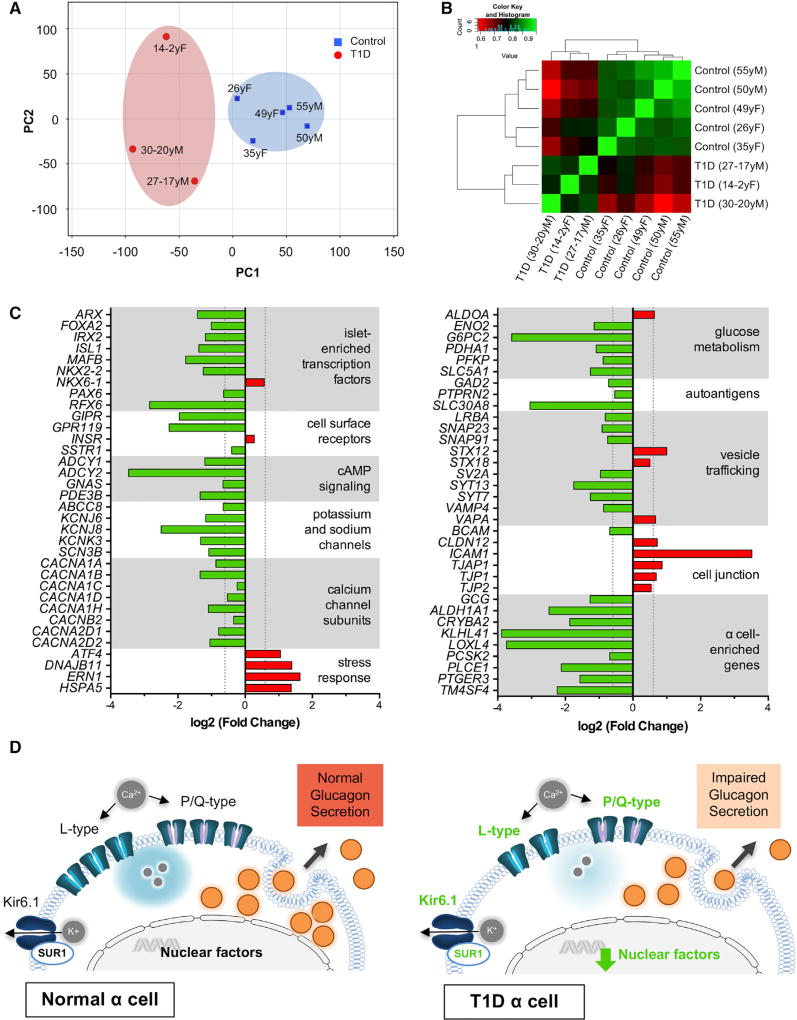
Many patients with type 1 diabetes (T1D) have residual β cells producing small amounts of C-peptide long after disease onset but develop an inadequate glucagon response to hypoglycemia following T1D diagnosis. The features of these residual β cells and α cells in the islet endocrine compartment are largely unknown, due to the difficulty of comprehensive investigation. By studying the T1D pancreas and isolated islets, we show that remnant β cells appeared to maintain several aspects of regulated insulin secretion. However, the function of T1D α cells was markedly reduced, and these cells had alterations in transcription factors constituting α and β cell identity. In the native pancreas and after placing the T1D islets into a non-autoimmune, normoglycemic in vivo environment, there was no evidence of α-to-β cell conversion. These results suggest an explanation for the disordered T1D counterregulatory glucagon response to hypoglycemia.
Keywords: alpha cells; glucagon; human; insulin; pancreatic islet; type 1 diabetes.
Copyright © 2018 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Alkorta-Aranburu G, Sukhanova M, Carmody D, Hoffman T, Wysinger L, Keller-Ramey J, Li Z, Johnson AK, Kobiernicki F, Botes S, et al. Improved molecular diagnosis of patients with neonatal diabetes using a combined next-generation sequencing and MS-MLPA approach. J. Pediatr. Endocrinol. Metab. 2016;29:523–531. - PubMed
-
- Balamurugan AN, Chang Y, Fung JJ, Trucco M, Bottino R. Flexible management of enzymatic digestion improves human islet isolation outcome from sub-optimal donor pancreata. Am. J. Transplant. 2003;3:1135–1142. - PubMed
-
- Bolli G, de Feo P, Compagnucci P, Cartechini MG, Angeletti G, Santeusanio F, Brunetti P, Gerich JE. Abnormal glucose counterregulation in insulin-dependent diabetes mellitus. Interaction of anti-insulin antibodies and impaired glucagon and epinephrine secretion. Diabetes. 1983;32:134–141. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK090570/DK/NIDDK NIH HHS/United States
- UC4 DK104211/DK/NIDDK NIH HHS/United States
- P30 DK058404/DK/NIDDK NIH HHS/United States
- UC4 DK108120/DK/NIDDK NIH HHS/United States
- S10 OD021630/OD/NIH HHS/United States
- U01 DK072473/DK/NIDDK NIH HHS/United States
- UC4 DK112232/DK/NIDDK NIH HHS/United States
- U01 DK089572/DK/NIDDK NIH HHS/United States
- P30 CA068485/CA/NCI NIH HHS/United States
- F30 DK118830/DK/NIDDK NIH HHS/United States
- R01 DK097829/DK/NIDDK NIH HHS/United States
- R21 DK066636/DK/NIDDK NIH HHS/United States
- T32 GM007347/GM/NIGMS NIH HHS/United States
- I01 BX000666/BX/BLRD VA/United States
- R01 DK094199/DK/NIDDK NIH HHS/United States
- P30 DK020593/DK/NIDDK NIH HHS/United States
- R24 DK106755/DK/NIDDK NIH HHS/United States
- R01 DK050203/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
