

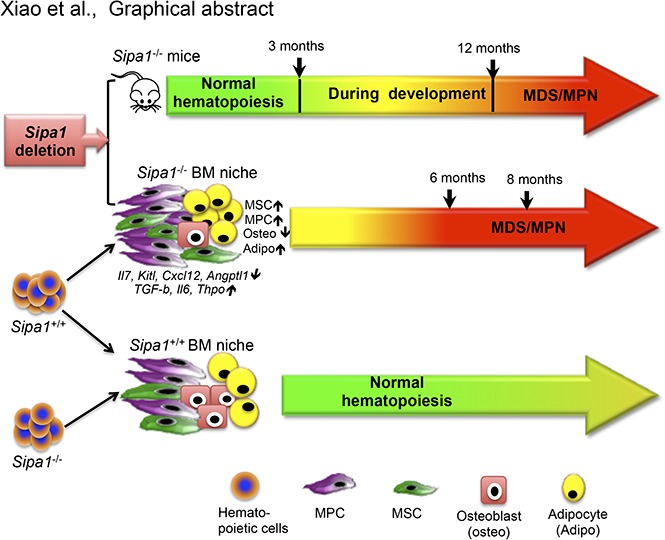
Sipa1 deficiency-induced bone marrow niche alterations lead to the initiation of myeloproliferative neoplasm
- PMID: 29514790
- PMCID: PMC5851419
- DOI: 10.1182/bloodadvances.2017013599
Sipa1 deficiency-induced bone marrow niche alterations lead to the initiation of myeloproliferative neoplasm
Abstract
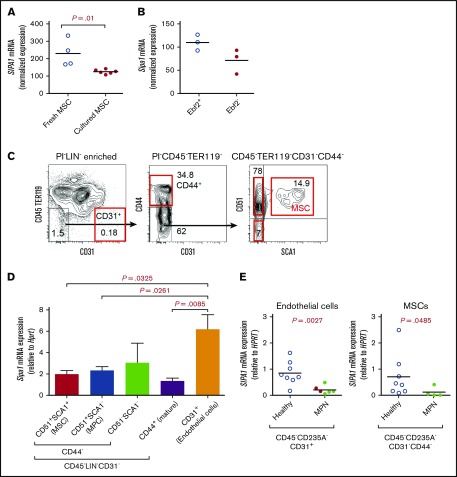
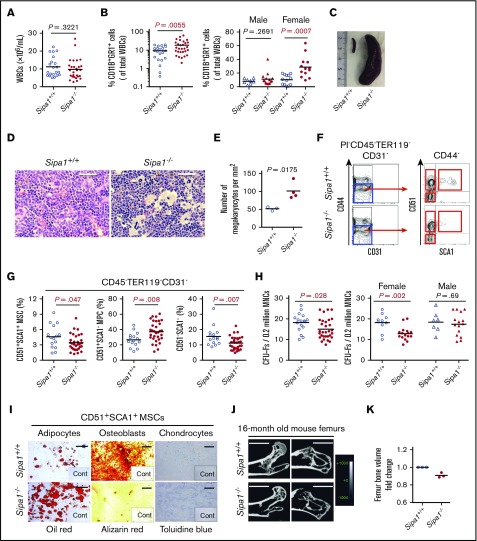
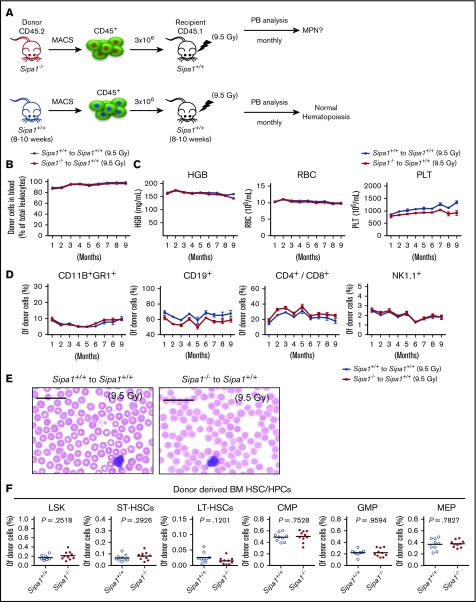
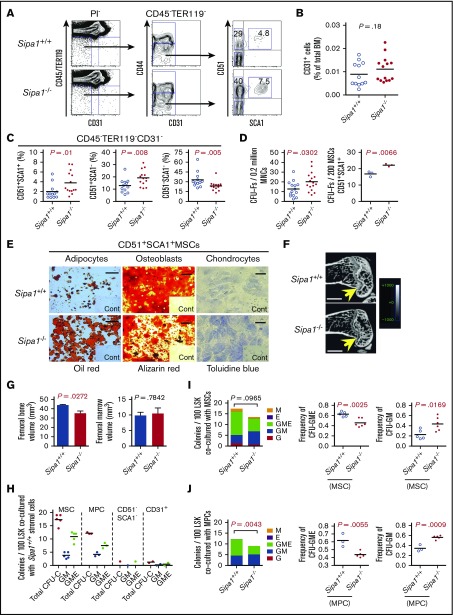
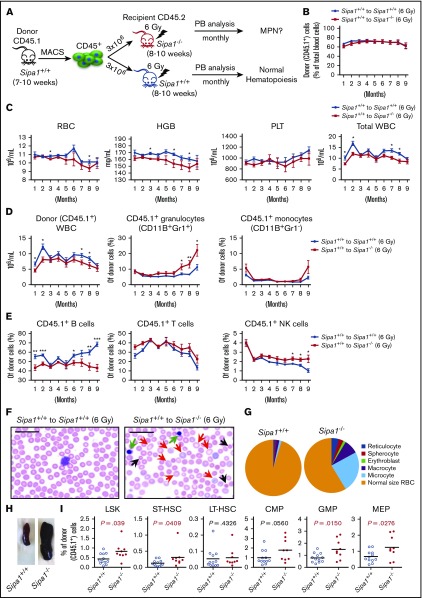
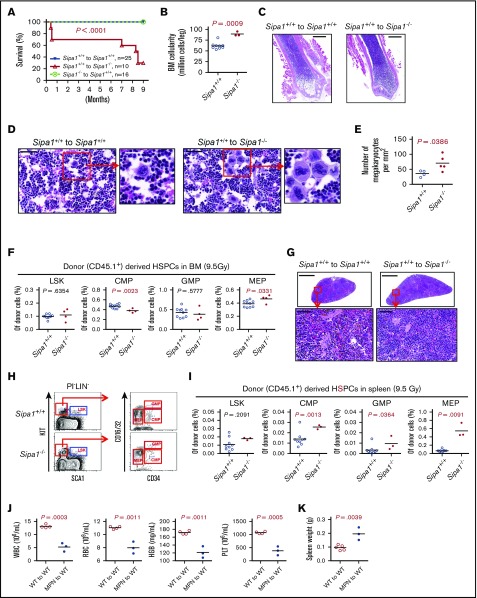
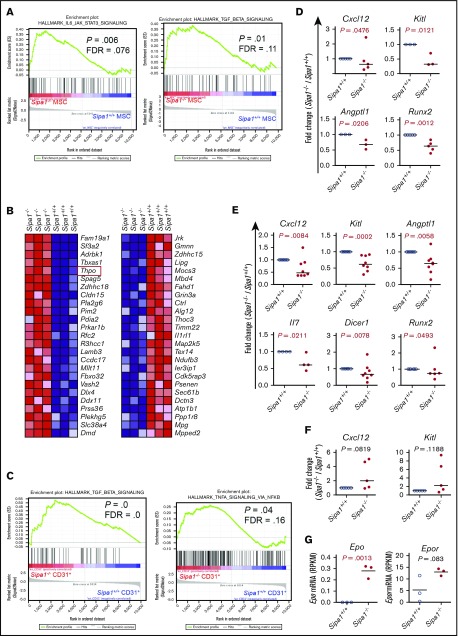
Mutations of signal-induced proliferation-associated gene 1 (SIPA1), a RAP1 GTPase-activating protein, were reported in patients with juvenile myelomonocytic leukemia, a childhood myelodysplastic/myeloproliferative neoplasm (MDS/MPN). Sipa1 deficiency in mice leads to the development of age-dependent MPN. However, Sipa1 expression in bone marrow (BM) microenvironment and its effect on the pathogenesis of MPN remain unclear. We here report that Sipa1 is expressed in human and mouse BM stromal cells and downregulated in these cells from patients with MPN or MDS/MPN at diagnosis. By using the Sipa1-/- MPN mouse model, we find that Sipa1 deletion causes phenotypic and functional alterations of BM mesenchymal stem and progenitor cells prior to the initiation of the MPN. Importantly, the altered Sipa1-/- BM niche is required for the development of MDS/MPN following transplantation of normal hematopoietic cells. RNA sequencing reveals an enhanced inflammatory cytokine signaling and dysregulated Dicer1, Kitl, Angptl1, Cxcl12, and Thpo in the Sipa1-/- BM cellular niches. Our data suggest that Sipa1 expression in the BM niche is critical for maintaining BM niche homeostasis. Moreover, Sipa1 loss-induced BM niche alterations likely enable evolution of clonal hematopoiesis to the hematological malignancies. Therefore, restoring Sipa1 expression or modulating the altered signaling pathways involved might offer therapeutic potential for MPN.
© 2018 by The American Society of Hematology.
Conflict of interest statement
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Figures
Similar articles
-
Distinct roles of mesenchymal stem and progenitor cells during the development of acute myeloid leukemia in mice.Blood Adv. 2018 Jun 26;2(12):1480-1494. doi: 10.1182/bloodadvances.2017013870. Blood Adv. 2018. PMID: 29945938 Free PMC article.
-
Sipa1 deficiency unleashes a host-immune mechanism eradicating chronic myelogenous leukemia-initiating cells.Nat Commun. 2018 Mar 2;9(1):914. doi: 10.1038/s41467-018-03307-8. Nat Commun. 2018. PMID: 29500416 Free PMC article.
-
Leukaemogenic effects of Ptpn11 activating mutations in the stem cell microenvironment.Nature. 2016 Nov 10;539(7628):304-308. doi: 10.1038/nature20131. Epub 2016 Oct 26. Nature. 2016. PMID: 27783593 Free PMC article.
-
Bone marrow niche in the myelodysplastic syndromes.Leuk Res. 2015 Oct;39(10):1020-7. doi: 10.1016/j.leukres.2015.06.017. Epub 2015 Jul 14. Leuk Res. 2015. PMID: 26276090 Review.
-
Mesenchymal Stem and Progenitor Cells in Normal and Dysplastic Hematopoiesis-Masters of Survival and Clonality?Int J Mol Sci. 2016 Jun 27;17(7):1009. doi: 10.3390/ijms17071009. Int J Mol Sci. 2016. PMID: 27355944 Free PMC article. Review.
Cited by
-
Chromosomal abnormalities of mesenchymal stromal cells in hematological malignancies.Oncogene. 2025 Sep;44(35):3155-3170. doi: 10.1038/s41388-025-03528-4. Epub 2025 Aug 14. Oncogene. 2025. PMID: 40813803 Free PMC article. Review.
-
Hematopoietic Stem Cell Niches and Signals Controlling Immune Cell Development and Maintenance of Immunological Memory.Front Immunol. 2020 Nov 26;11:600127. doi: 10.3389/fimmu.2020.600127. eCollection 2020. Front Immunol. 2020. PMID: 33324418 Free PMC article. Review.
-
Influence of the Bone Marrow Microenvironment on Hematopoietic Stem Cell Behavior Post-Allogeneic Transplantation: Development of Clonal Hematopoiesis and Telomere Dynamics.Int J Mol Sci. 2024 Sep 24;25(19):10258. doi: 10.3390/ijms251910258. Int J Mol Sci. 2024. PMID: 39408588 Free PMC article.
-
A senescence stress secretome is a hallmark of therapy-related myeloid neoplasm stromal tissue occurring soon after cytotoxic exposure.Leukemia. 2022 Nov;36(11):2678-2689. doi: 10.1038/s41375-022-01686-y. Epub 2022 Aug 29. Leukemia. 2022. PMID: 36038666 Free PMC article.
-
The Instructive Role of the Bone Marrow Niche in Aging and Leukemia.Curr Stem Cell Rep. 2018;4(4):291-298. doi: 10.1007/s40778-018-0143-7. Epub 2018 Oct 12. Curr Stem Cell Rep. 2018. PMID: 30524938 Free PMC article. Review.
References
-
- Schofield R. The relationship between the spleen colony-forming cell and the haemopoietic stem cell. Blood Cells. 1978;4(1-2):7-25. - PubMed
-
- Hoggatt J, Kfoury Y, Scadden DT. Hematopoietic stem cell niche in health and disease. Annu Rev Pathol. 2016;11:555-581. - PubMed
-
- Pittenger MF, Mackay AM, Beck SC, et al. . Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284(5411):143-147. - PubMed
-
- Omatsu Y, Sugiyama T, Kohara H, et al. . The essential functions of adipo-osteogenic progenitors as the hematopoietic stem and progenitor cell niche. Immunity. 2010;33(3):387-399. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
