

Spatiotemporal compartmentalization of hepatic NADH and NADPH metabolism
- PMID: 29514978
- PMCID: PMC5961030
- DOI: 10.1074/jbc.TM117.000258
Spatiotemporal compartmentalization of hepatic NADH and NADPH metabolism
Abstract
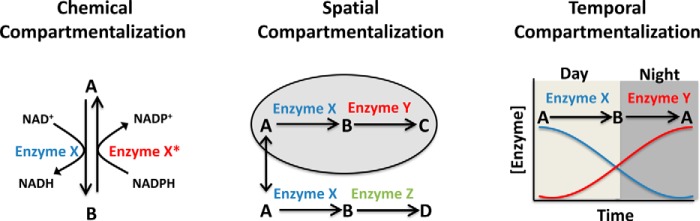
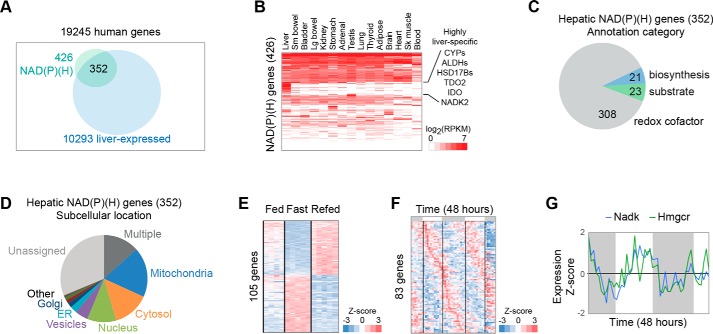
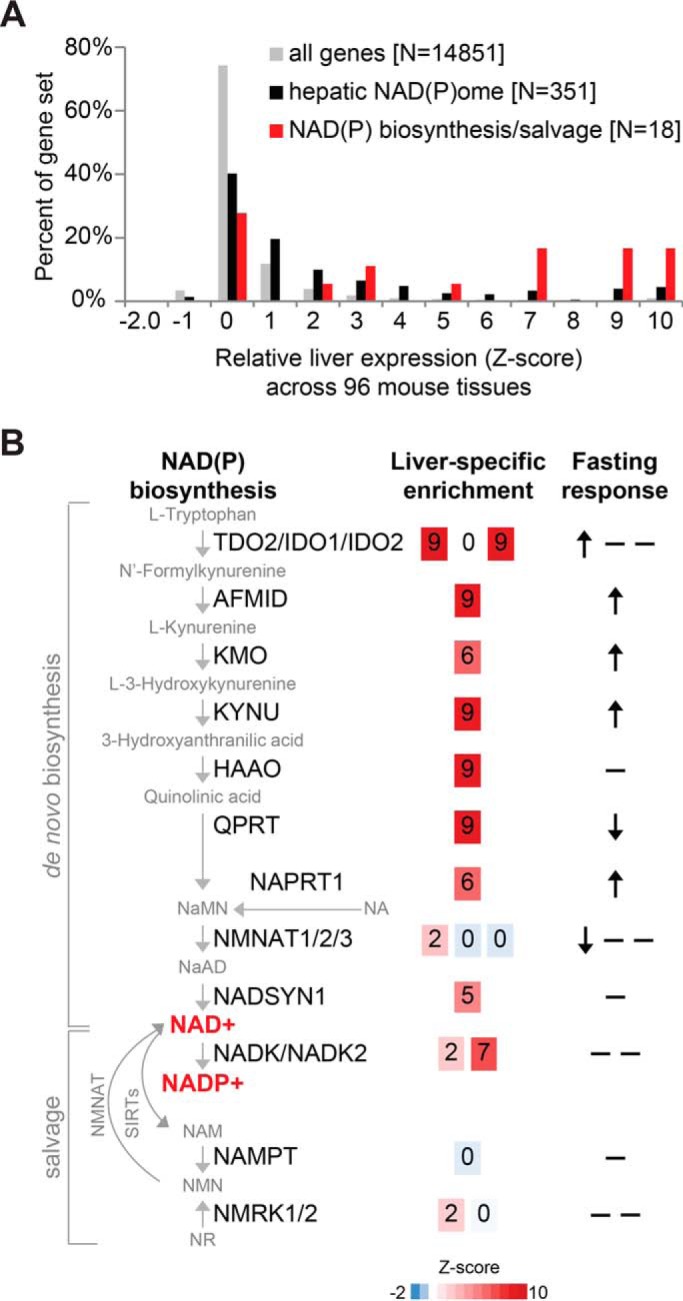
Compartmentalization is a fundamental design principle of eukaryotic metabolism. Here, we review the compartmentalization of NAD+/NADH and NADP+/NADPH with a focus on the liver, an organ that experiences the extremes of biochemical physiology each day. Historical studies of the liver, using classical biochemical fractionation and measurements of redox-coupled metabolites, have given rise to the prevailing view that mitochondrial NAD(H) pools tend to be oxidized and important for energy homeostasis, whereas cytosolic NADP(H) pools tend to be highly reduced for reductive biosynthesis. Despite this textbook view, many questions still remain as to the relative size of these subcellular pools and their redox ratios in different physiological states, and to what extent such redox ratios are simply indicators versus drivers of metabolism. By performing a bioinformatic survey, we find that the liver expresses 352 known or predicted enzymes composing the hepatic NAD(P)ome, i.e. the union of all predicted enzymes producing or consuming NADP(H) or NAD(H) or using them as a redox co-factor. Notably, less than half are predicted to be localized within the cytosol or mitochondria, and a very large fraction of these genes exhibit gene expression patterns that vary during the time of day or in response to fasting or feeding. A future challenge lies in applying emerging new genetic tools to measure and manipulate in vivo hepatic NADP(H) and NAD(H) with subcellular and temporal resolution. Insights from such fundamental studies will be crucial in deciphering the pathogenesis of very common diseases known to involve alterations in hepatic NAD(P)H, such as diabetes and fatty liver disease.
Keywords: NAD biosynthesis; NAD(P)ome; cell metabolism; hepatic metabolism; hepatocyte; intermediary metabolism; liver; liver metabolism; nicotinamide adenine dinucleotide (NAD); nicotinamide adenine dinucleotide (NADH); oxidation-reduction (redox).
© 2018 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures

References
-
- Takamura-Enya T., Watanabe M., Totsuka Y., Kanazawa T., Matsushima-Hibiya Y., Koyama K., Sugimura T., and Wakabayashi K. (2001) Mono(ADP-ribosyl)ation of 2′-deoxyguanosine residue in DNA by an apoptosis-inducing protein, pierisin-1, from cabbage butterfly. Proc. Natl. Acad. Sci. U.S.A. 98, 12414–12419 10.1073/pnas.221444598 - DOI - PMC - PubMed
-
- Hirschey M. D., Shimazu T., Goetzman E., Jing E., Schwer B., Lombard D. B., Grueter C. A., Harris C., Biddinger S., Ilkayeva O. R., Stevens R. D., Li Y., Saha A. K., Ruderman N. B., Bain J. R., Newgard C. B., et al. (2010) SIRT3 regulates mitochondrial fatty-acid oxidation by reversible enzyme deacetylation. Nature 464, 121–125 10.1038/nature08778 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
