

Peripheral tissues reprogram CD8+ T cells for pathogenicity during graft-versus-host disease
- PMID: 29515032
- PMCID: PMC5922296
- DOI: 10.1172/jci.insight.97011
Peripheral tissues reprogram CD8+ T cells for pathogenicity during graft-versus-host disease
Abstract
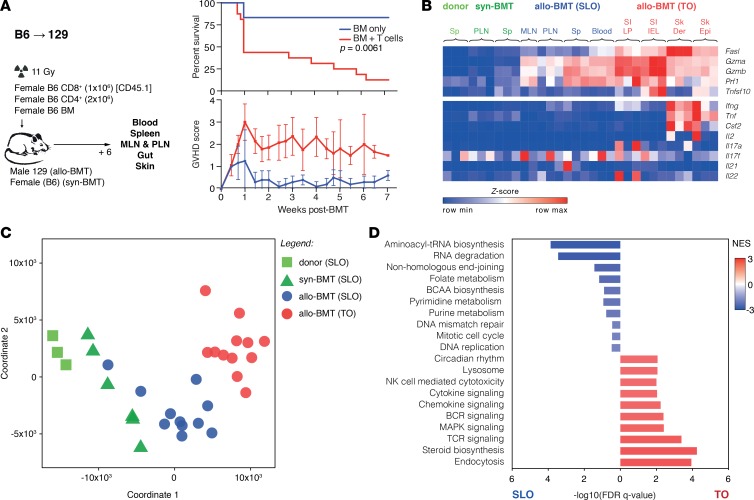
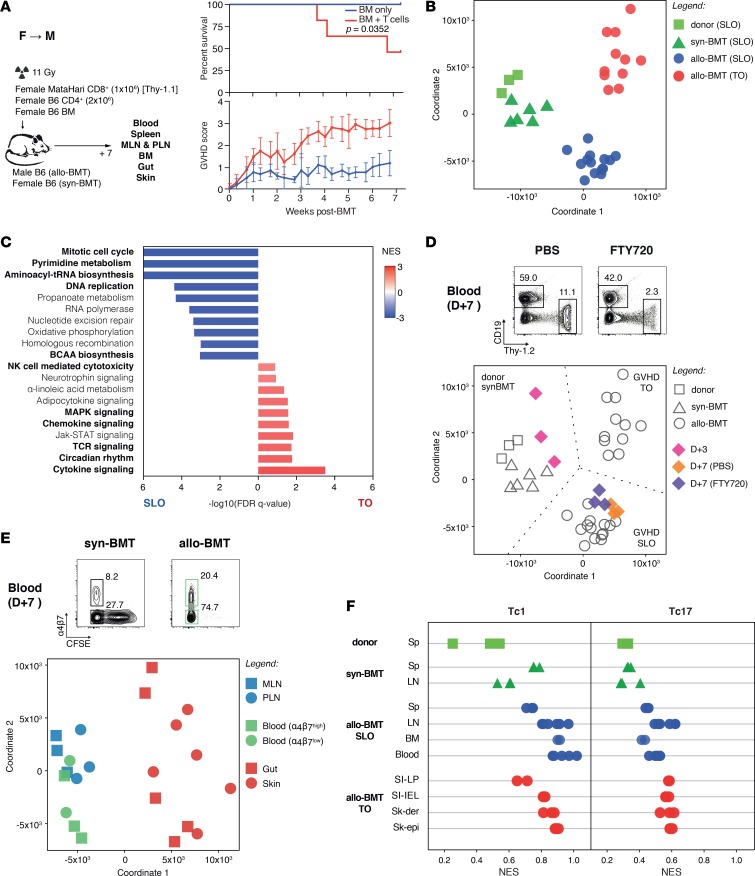
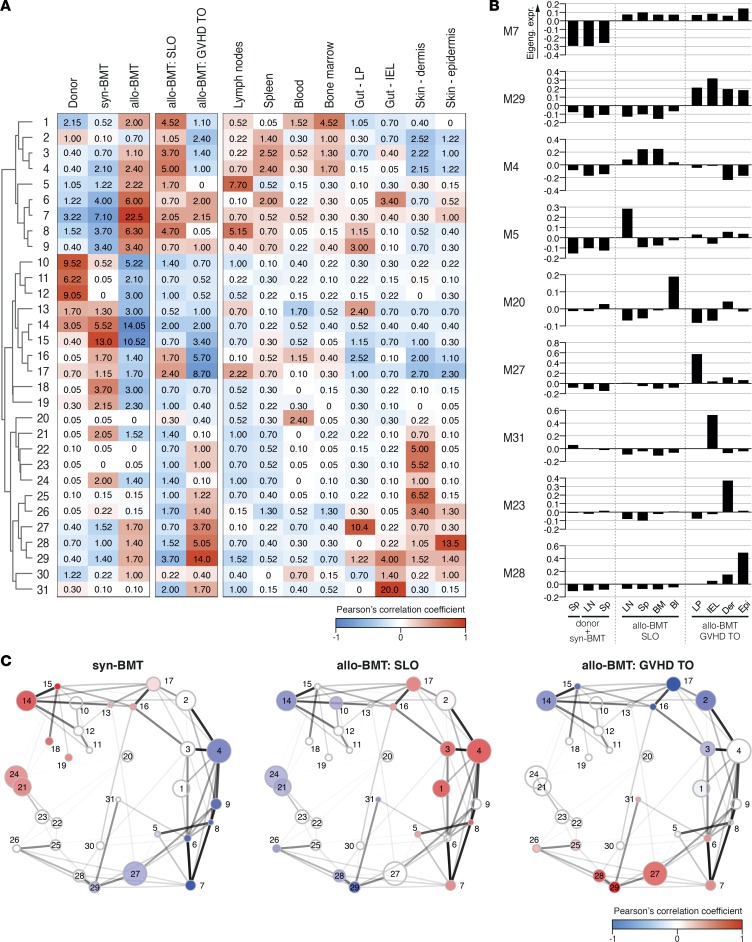
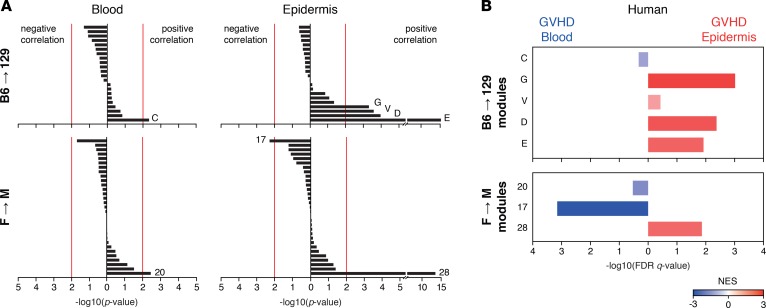
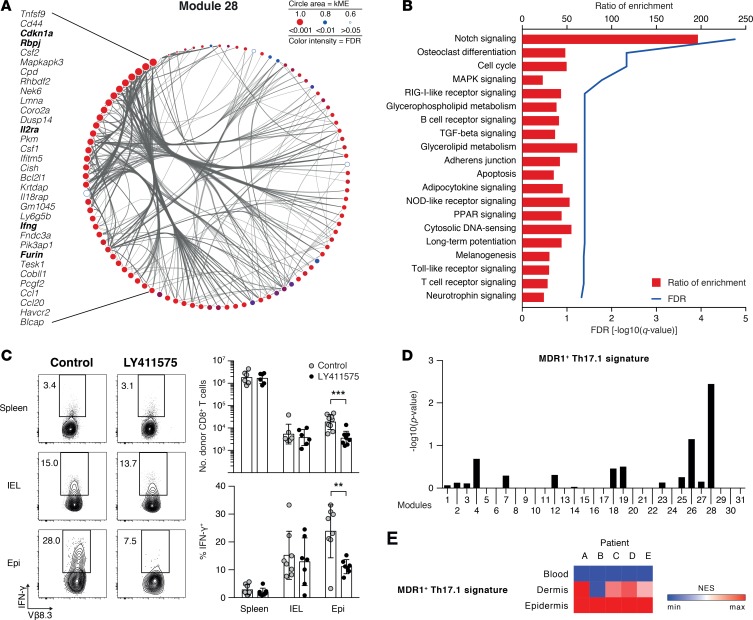
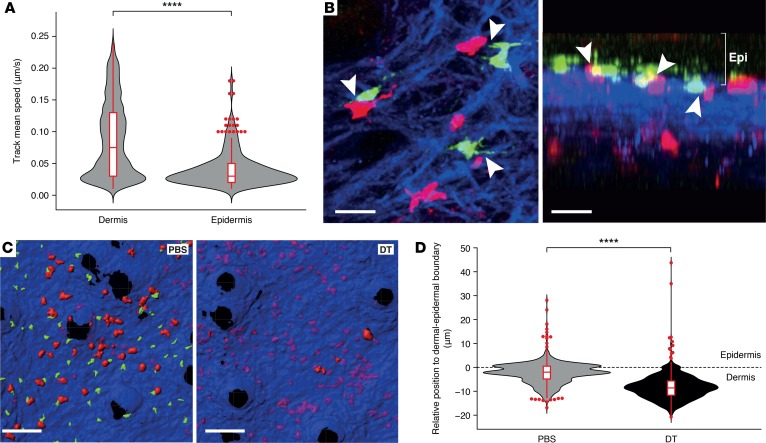
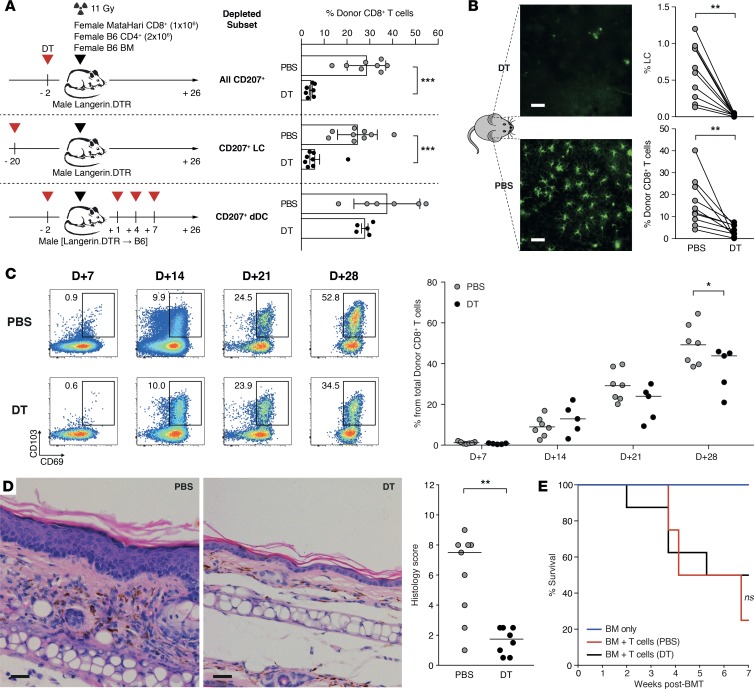
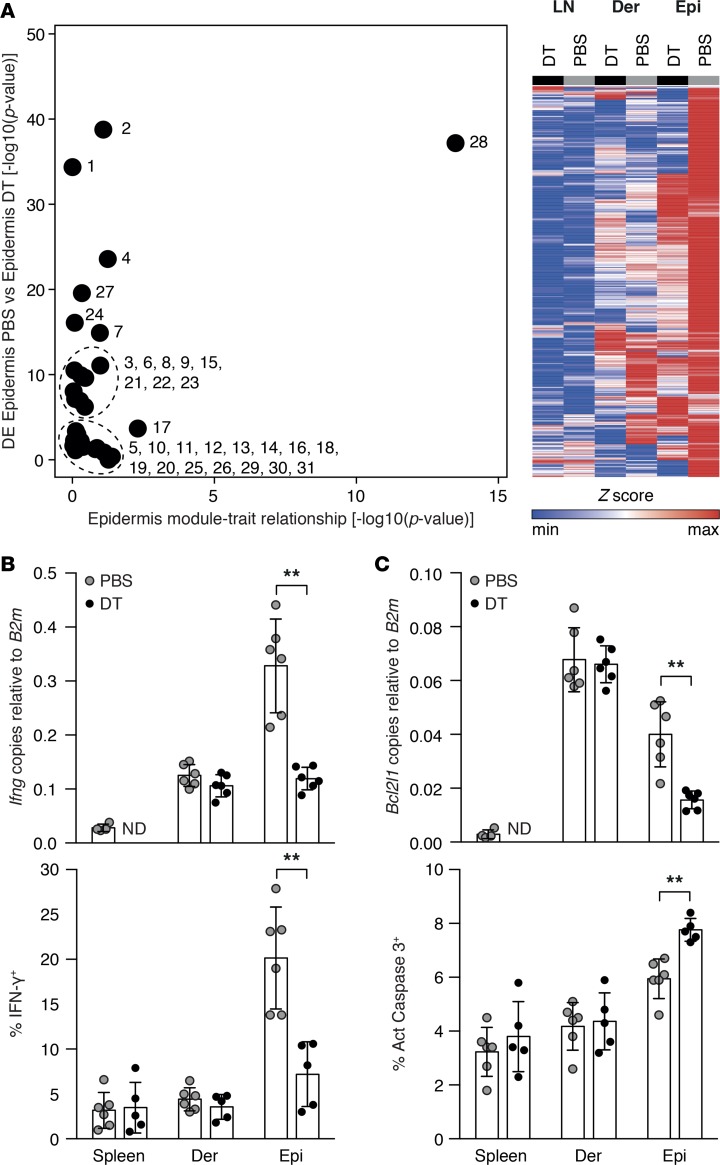
Graft-versus-host disease (GVHD) is a life-threatening complication of allogeneic stem cell transplantation induced by the influx of donor-derived effector T cells (TE) into peripheral tissues. Current treatment strategies rely on targeting systemic T cells; however, the precise location and nature of instructions that program TE to become pathogenic and trigger injury are unknown. We therefore used weighted gene coexpression network analysis to construct an unbiased spatial map of TE differentiation during the evolution of GVHD and identified wide variation in effector programs in mice and humans according to location. Idiosyncrasy of effector programming in affected organs did not result from variation in T cell receptor repertoire or the selection of optimally activated TE. Instead, TE were reprogrammed by tissue-autonomous mechanisms in target organs for site-specific proinflammatory functions that were highly divergent from those primed in lymph nodes. In the skin, we combined the correlation-based network with a module-based differential expression analysis and showed that Langerhans cells provided in situ instructions for a Notch-dependent T cell gene cluster critical for triggering local injury. Thus, the principal determinant of TE pathogenicity in GVHD is the final destination, highlighting the need for target organ-specific approaches to block immunopathology while avoiding global immune suppression.
Keywords: Immunology; Molecular pathology; Stem cell transplantation; T cell development; Transplantation.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
