

Sex differences in the regulation of social and anxiety-related behaviors: insights from vasopressin and oxytocin brain systems
- PMID: 29518698
- PMCID: PMC6055524
- DOI: 10.1016/j.conb.2018.02.011
Sex differences in the regulation of social and anxiety-related behaviors: insights from vasopressin and oxytocin brain systems
Abstract
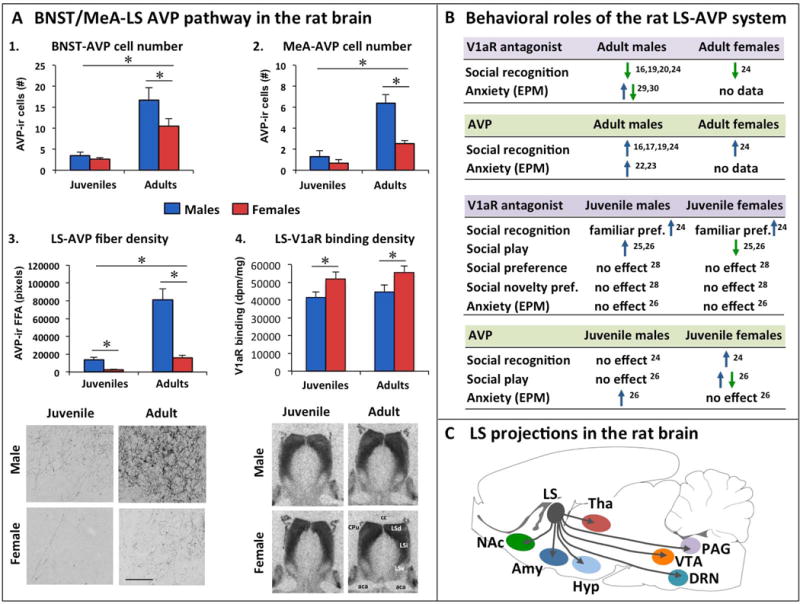
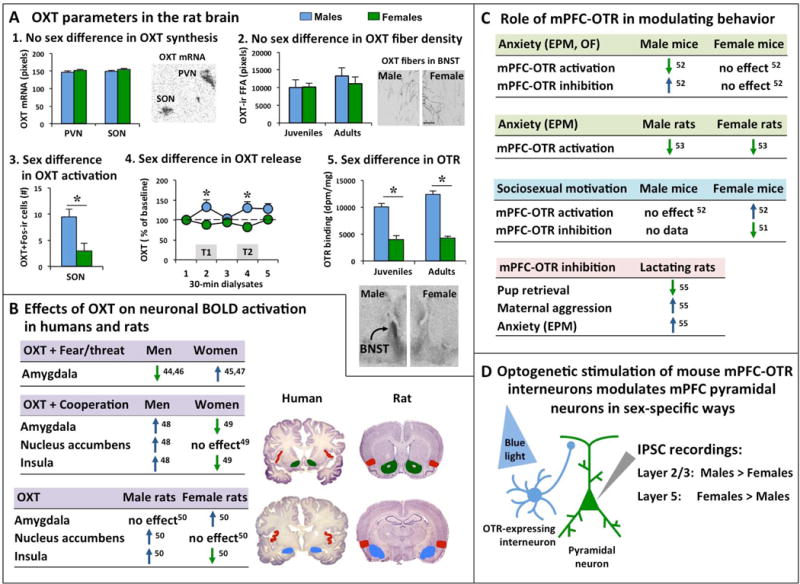
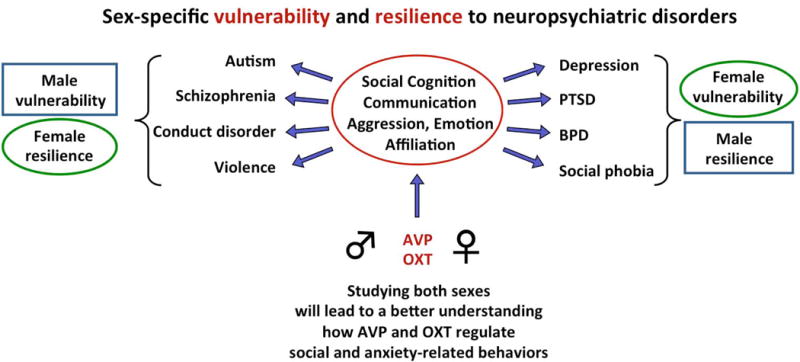
To understand how the brain regulates behavior, many variables must be taken into account, with sex as a prominent variable. In this review, we will discuss recent human and rodent studies showing the sex-specific involvement of the neuropeptides vasopressin and oxytocin in social and anxiety-related behaviors. We discuss that sex differences can be evident at pre-pubertal ages as seen in the sex-specific regulation of social recognition, social play, and anxiety by the vasopressin system in juvenile rats. We further discuss that the oxytocin system in humans and rodents alters brain activation, anxiety, and sociosexual motivation in sex-specific ways. Finally, we propose that knowledge of vasopressin and oxytocin mediated sex-specific brain mechanisms can provide essential insights into how these neuropeptide systems contribute to sex-specific vulnerability as well as resilience to perturbations, with subsequent relevance to social and emotional disorders.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
