

Structural basis for coupling protein transport and N-glycosylation at the mammalian endoplasmic reticulum
- PMID: 29519914
- PMCID: PMC6319373
- DOI: 10.1126/science.aar7899
Structural basis for coupling protein transport and N-glycosylation at the mammalian endoplasmic reticulum
Abstract
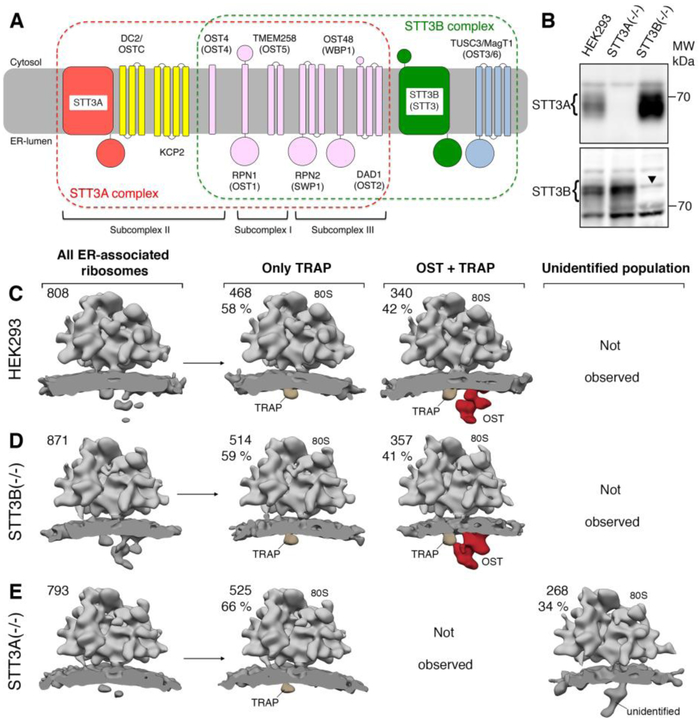
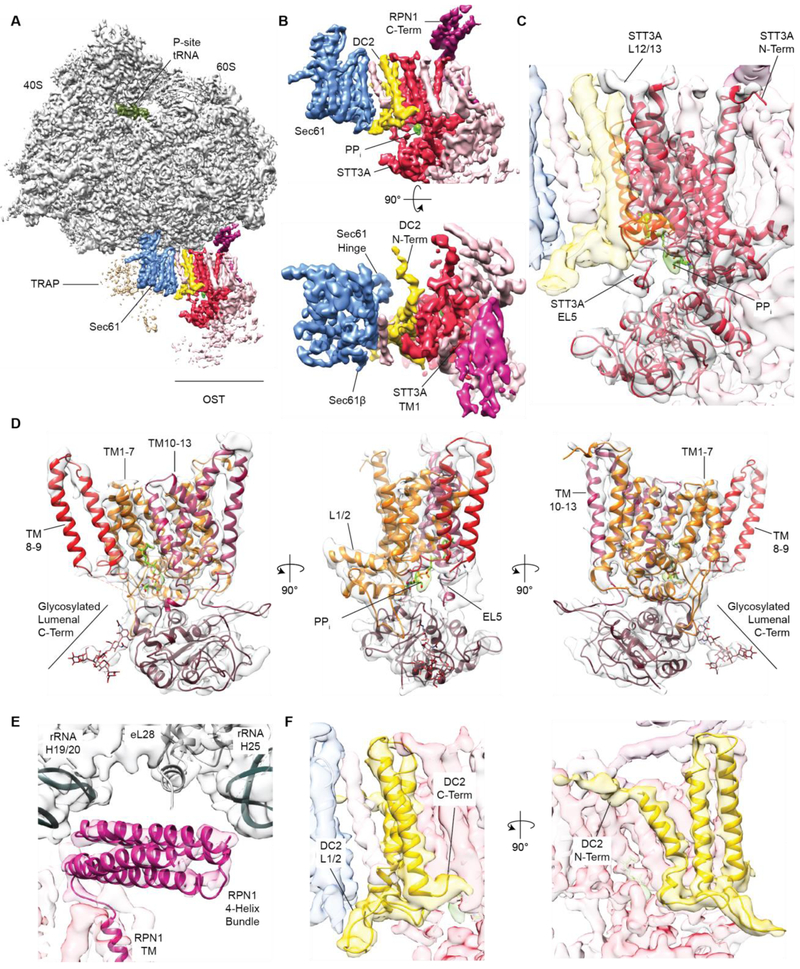
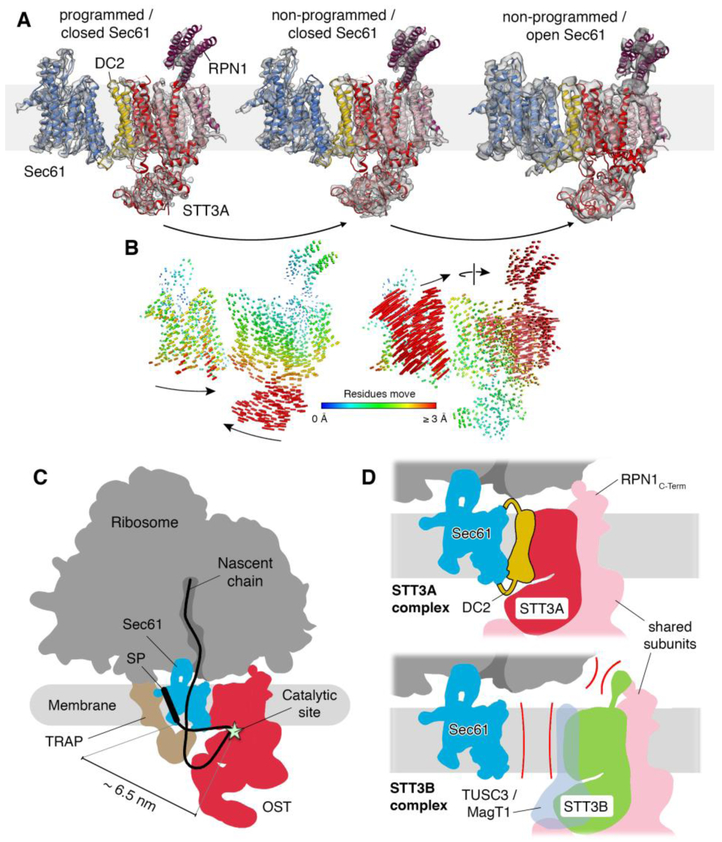
Protein synthesis, transport, and N-glycosylation are coupled at the mammalian endoplasmic reticulum by complex formation of a ribosome, the Sec61 protein-conducting channel, and oligosaccharyltransferase (OST). Here we used different cryo-electron microscopy approaches to determine structures of native and solubilized ribosome-Sec61-OST complexes. A molecular model for the catalytic OST subunit STT3A (staurosporine and temperature sensitive 3A) revealed how it is integrated into the OST and how STT3-paralog specificity for translocon-associated OST is achieved. The OST subunit DC2 was placed at the interface between Sec61 and STT3A, where it acts as a versatile module for recruitment of STT3A-containing OST to the ribosome-Sec61 complex. This detailed structural view on the molecular architecture of the cotranslational machinery for N-glycosylation provides the basis for a mechanistic understanding of glycoprotein biogenesis at the endoplasmic reticulum.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Competing interests:
We declare no competing interests.
Figures
References
-
- Pfeffer S, Dudek J, Zimmermann R, Forster F, Organization of the native ribosome-translocon complex at the mammalian endoplasmic reticulum membrane. Biochim Biophys Acta 1860, 2122 (October, 2016). - PubMed
-
- Voorhees RM, Hegde RS, Toward a structural understanding of co-translational protein translocation. Curr Opin Cell Biol 41, 91 (August, 2016). - PubMed
-
- Rapoport TA, Li L, Park E, Structural and Mechanistic Insights into Protein Translocation. Annu. Rev. Cell Dev. Biol 33, 369 (October 6, 2017). - PubMed
-
- Beckmann R et al., Alignment of conduits for the nascent polypeptide chain in the ribosome-Sec61 complex. Science 278, 2123 (December 19, 1997). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
