

Mice with reduced DAT levels recreate seasonal-induced switching between states in bipolar disorder
- PMID: 29520059
- PMCID: PMC6006292
- DOI: 10.1038/s41386-018-0031-y
Mice with reduced DAT levels recreate seasonal-induced switching between states in bipolar disorder
Abstract
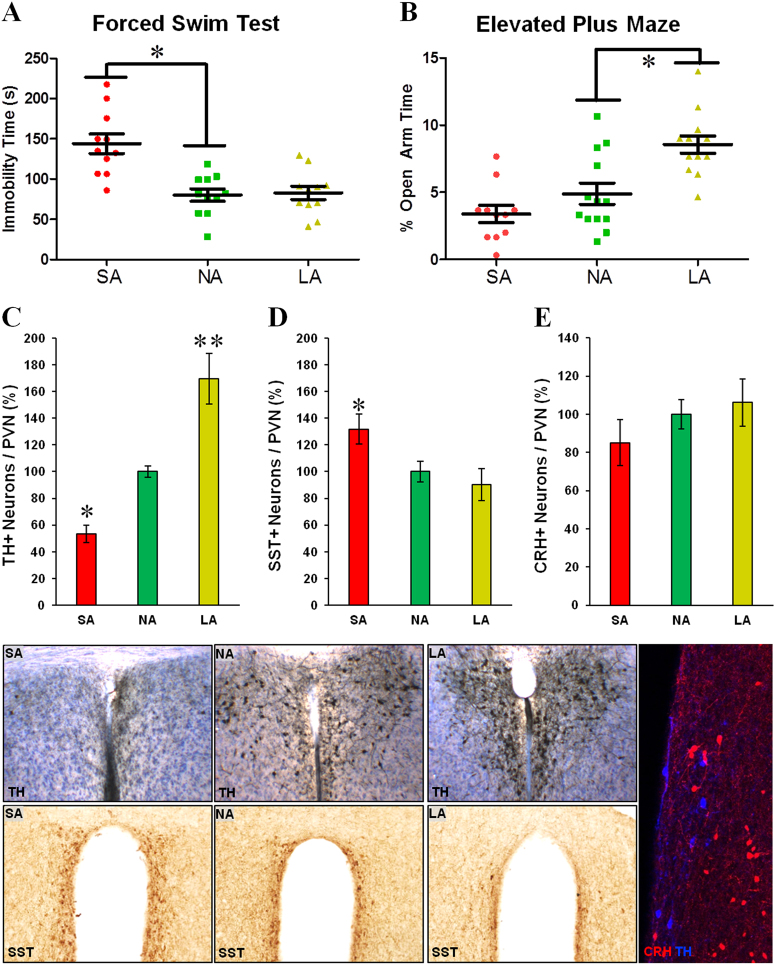
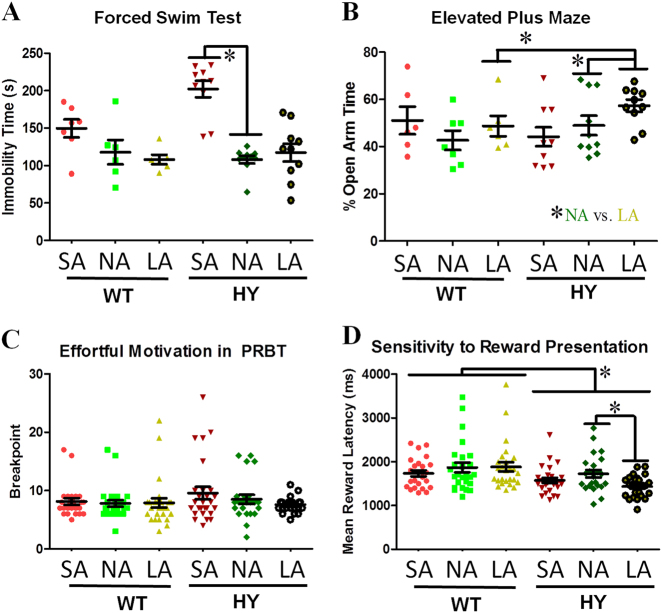
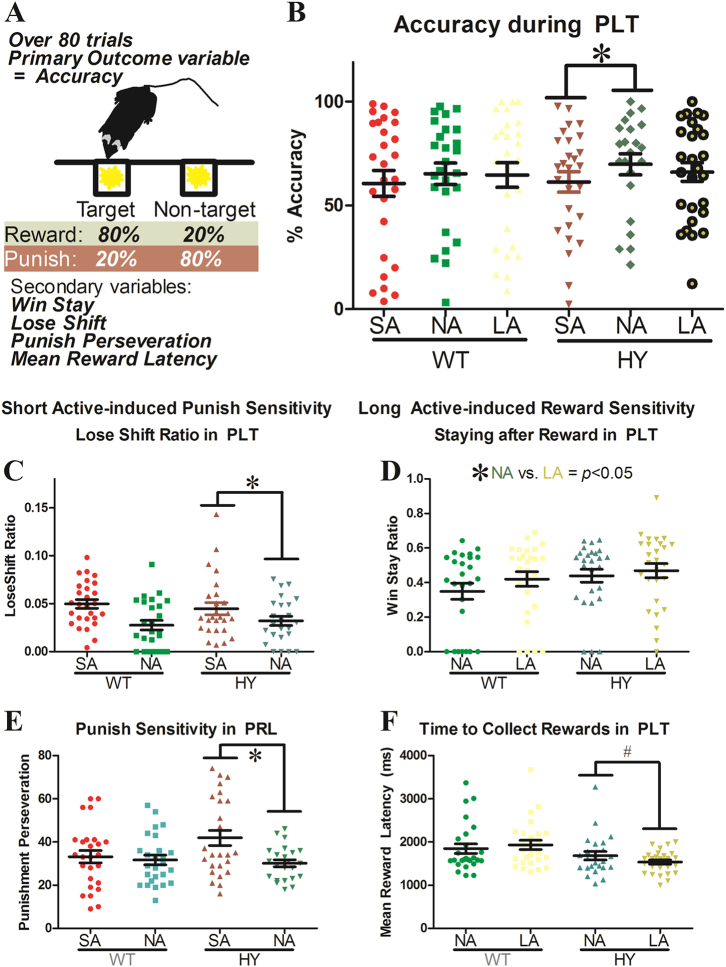
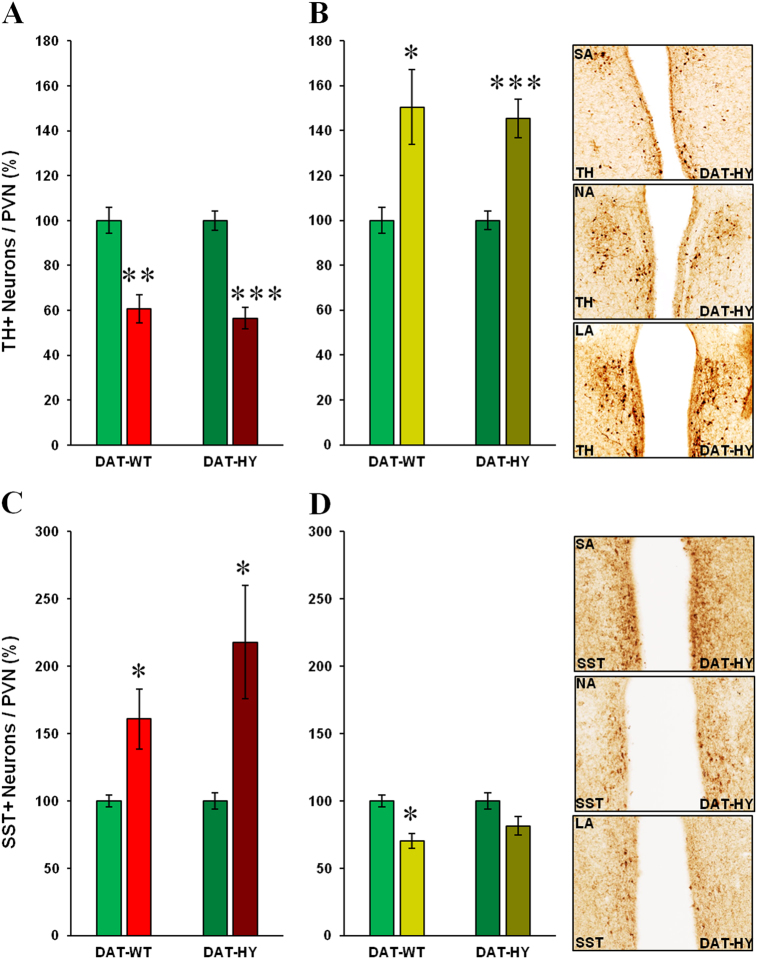
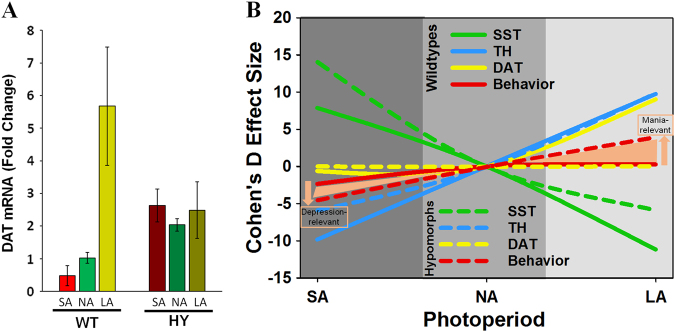
Developing novel therapeutics for bipolar disorder (BD) has been hampered by limited mechanistic knowledge how sufferers switch between mania and depression-how the same brain can switch between extreme states-described as the "holy grail" of BD research. Strong evidence implicates seasonally-induced switching between states, with mania associated with summer-onset, depression with winter-onset. Determining mechanisms of and sensitivity to such switching is required. C57BL/6J and dopamine transporter hypomorphic (DAT-HY 50% expression) mice performed a battery of psychiatry-relevant behavioral tasks following 2-week housing in chambers under seasonally relevant photoperiod extremes. Summer-like and winter-like photoperiod exposure induced mania-relevant and depression-relevant behaviors respectively in mice. This behavioral switch paralleled neurotransmitter switching from dopamine to somatostatin in hypothalamic neurons (receiving direct input from the photoperiod-processing center, the suprachiasmatic nucleus). Mice with reduced DAT expression exhibited hypersensitivity to these summer-like and winter-like photoperiods, including more extreme mania-relevant (including reward sensitivity during reinforcement learning), and depression-relevant (including punishment-sensitivity and loss-sensitivity during reinforcement learning) behaviors. DAT mRNA levels switched in wildtype littermate mice across photoperiods, an effect not replicated in DAT hypomorphic mice. This inability to adjust DAT levels to match photoperiod-induced neurotransmitter switching as a homeostatic control likely contributes to the susceptibility of DAT hypormophic mice to these switching photoperiods. These data reveal the potential contribution of photoperiod-induced neuroplasticity within an identified circuit of the hypothalamus, linked with reduced DAT function, underlying switching between states in BD. Further investigations of the circuit will likely identify novel therapeutic targets to block switching between states.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Comment in
-
Evidence for light-entrainment-induced switching between depression- & mania-relevant behaviors in mice.Neuropsychopharmacology. 2019 Sep;44(10):1679-1680. doi: 10.1038/s41386-019-0338-3. Epub 2019 Feb 20. Neuropsychopharmacology. 2019. PMID: 30787425 Free PMC article. No abstract available.
-
Switching winter and summer photoperiods in an animal model of bipolar disorder.Neuropsychopharmacology. 2019 Sep;44(10):1677-1678. doi: 10.1038/s41386-019-0337-4. Epub 2019 Feb 20. Neuropsychopharmacology. 2019. PMID: 30787426 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
