

Genetic and metabolic regulation of Mycobacterium tuberculosis acid growth arrest
- PMID: 29520087
- PMCID: PMC5843633
- DOI: 10.1038/s41598-018-22343-4
Genetic and metabolic regulation of Mycobacterium tuberculosis acid growth arrest
Abstract
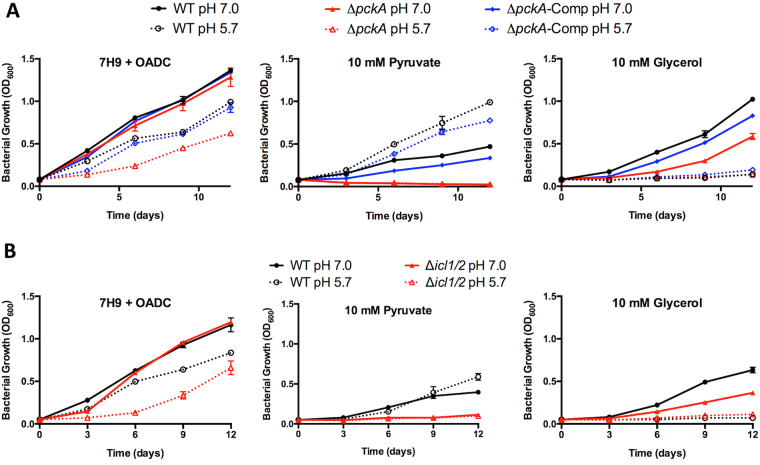
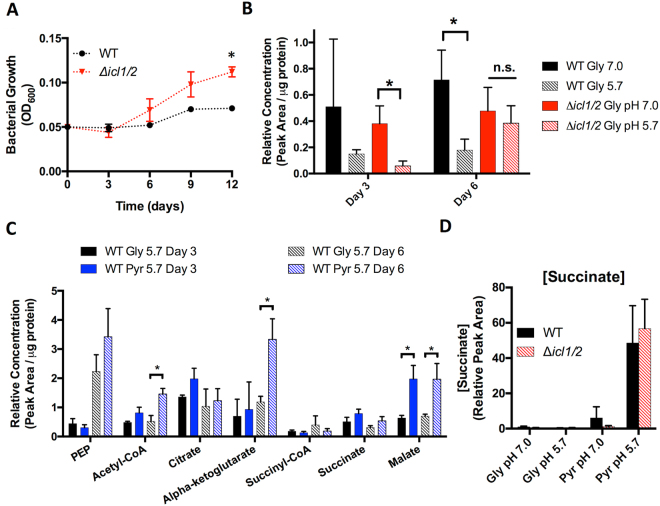
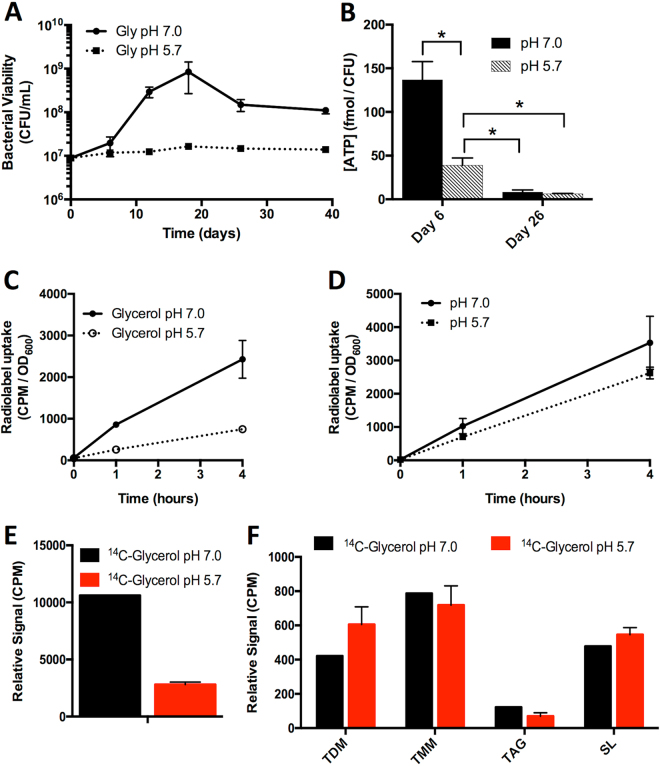
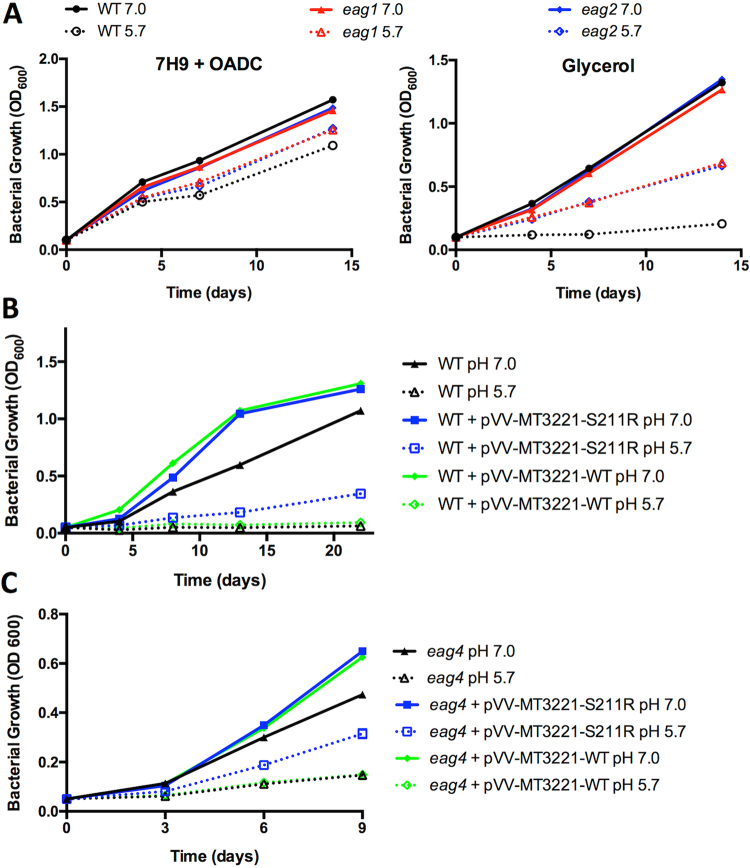
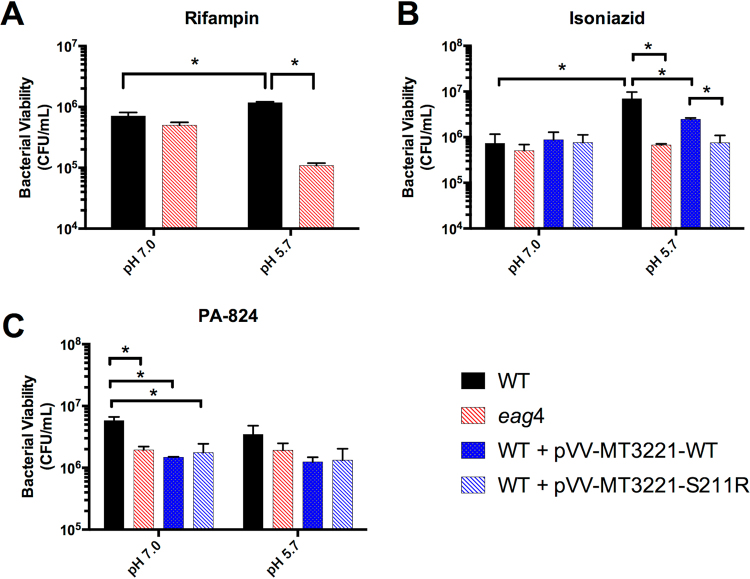
Mycobacterium tuberculosis (Mtb) senses and adapts to acidic environments during the course of infection. Acidic pH-dependent adaptations include the induction of metabolic genes associated with anaplerosis and growth arrest on specific carbon sources. Here we report that deletion of isocitrate lyase or phosphoenolpyruvate carboxykinase results in reduced growth at acidic pH and altered metabolite profiles, supporting that remodeling of anaplerotic metabolism is required for pH-dependent adaptation. Mtb cultured at pH 5.7 in minimal medium containing glycerol as a single carbon source exhibits an acid growth arrest phenotype, where the bacterium is non-replicating but viable and metabolically active. The bacterium assimilates and metabolizes glycerol and maintains ATP pools during acid growth arrest and becomes tolerant to detergent stress and the antibiotics isoniazid and rifampin. A forward genetic screen identified mutants that do not arrest their growth at acidic pH, including four enhanced acid growth (eag) mutants with three distinct mutations in the proline-proline-glutamate (PPE) gene MT3221 (also named ppe51). Overexpression of the MT3221(S211R) variant protein in wild type Mtb results in enhanced acid growth and reduced drug tolerance. These findings support that acid growth arrest is a genetically controlled, adaptive process and not simply a physiological limitation associated with acidic pH.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- WHO. Treatment of tuberculosis: guidelines (World Health Organization, 2010). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
