

Protein kinase C zeta suppresses low- or high-grade colorectal cancer (CRC) phenotypes by interphase centrosome anchoring
- PMID: 29520890
- PMCID: PMC5873423
- DOI: 10.1002/path.5035
Protein kinase C zeta suppresses low- or high-grade colorectal cancer (CRC) phenotypes by interphase centrosome anchoring
Abstract
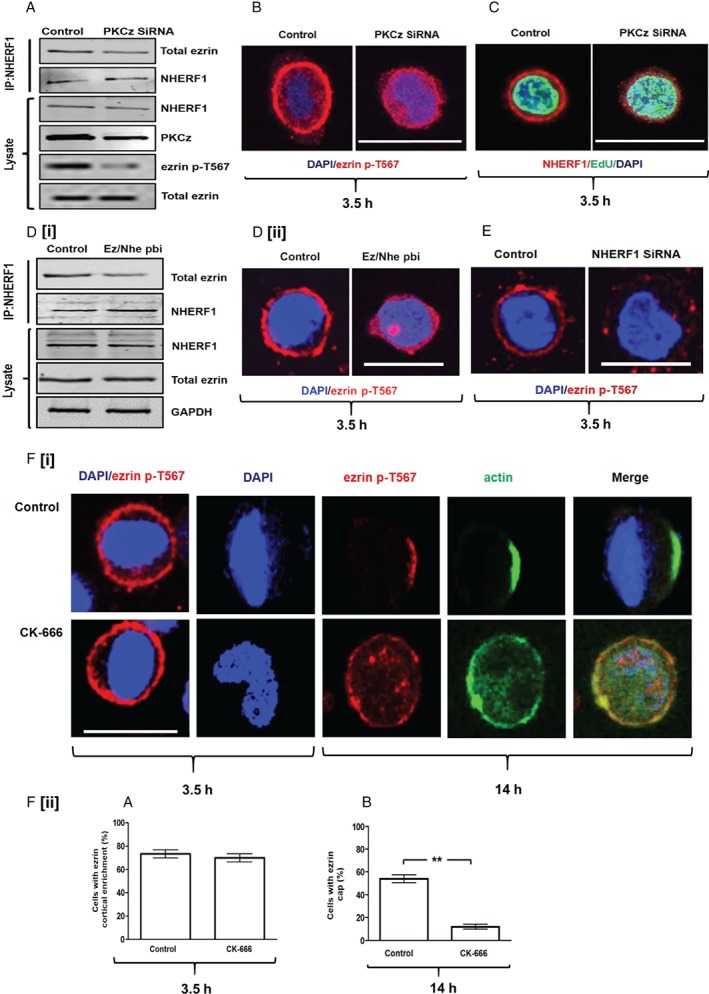
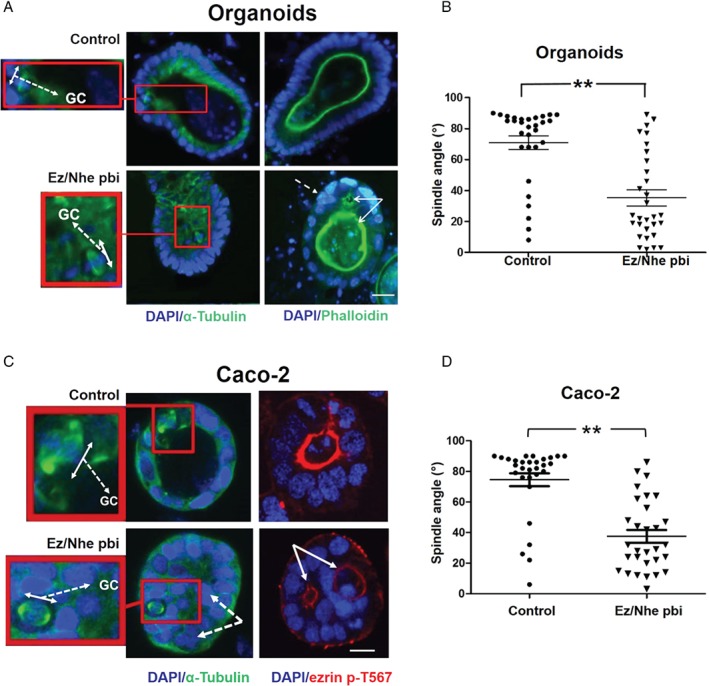
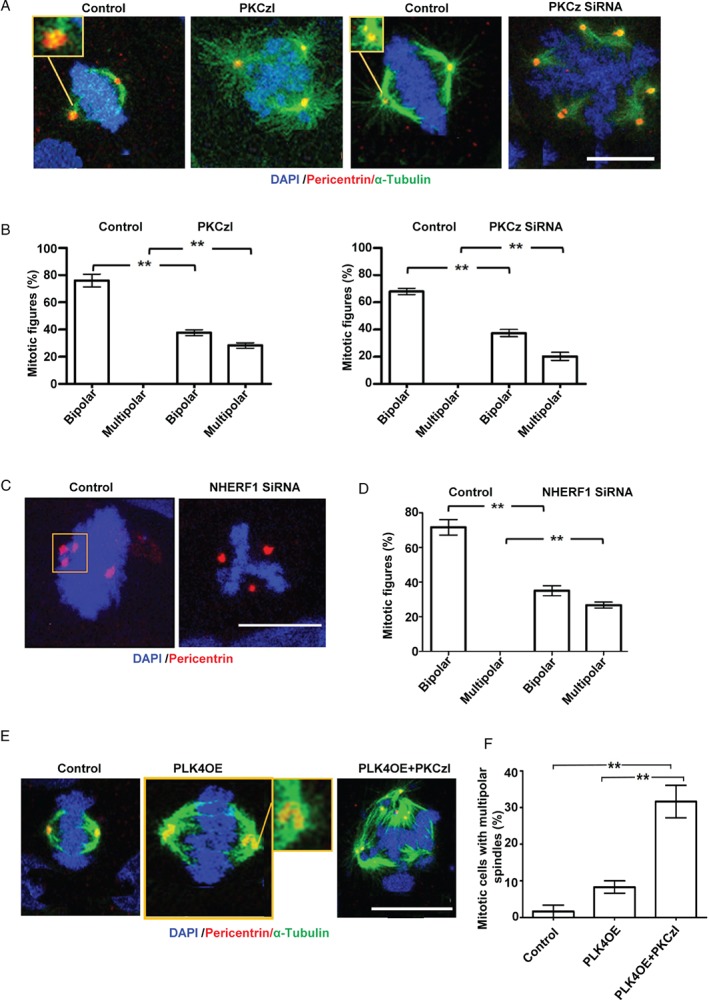
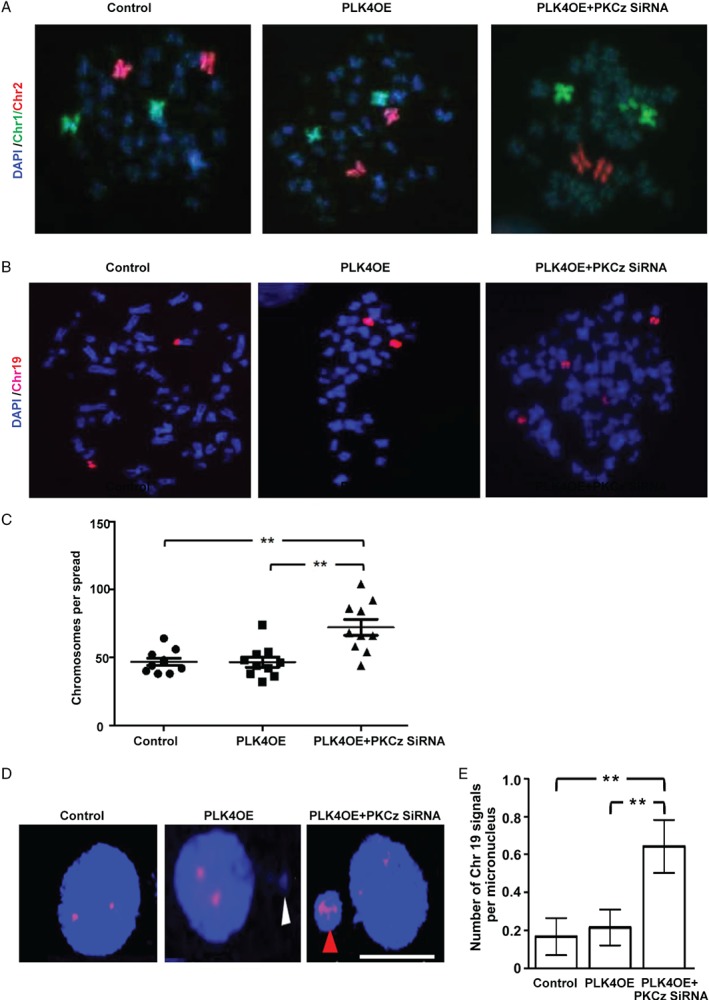
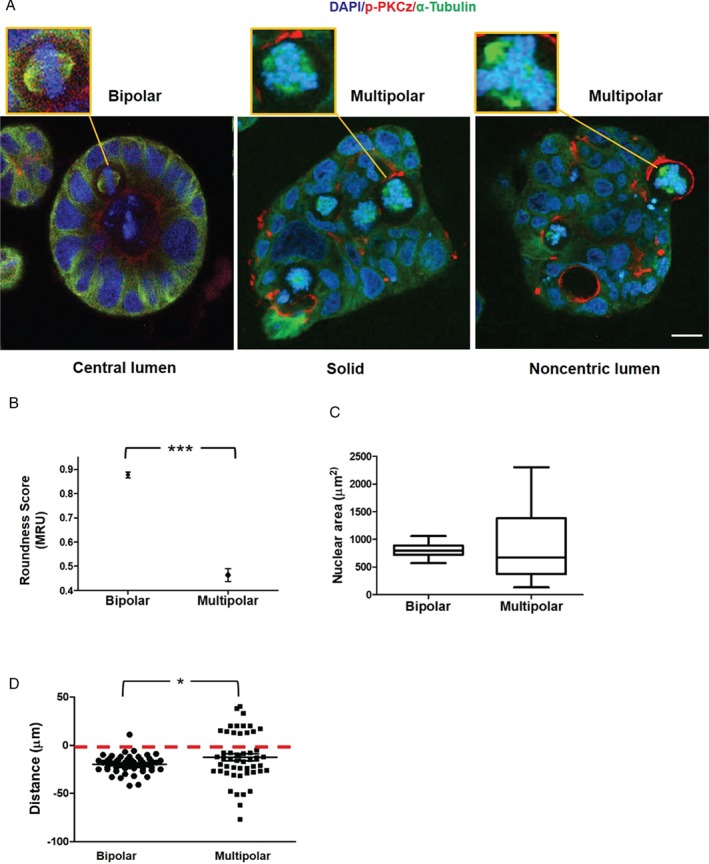
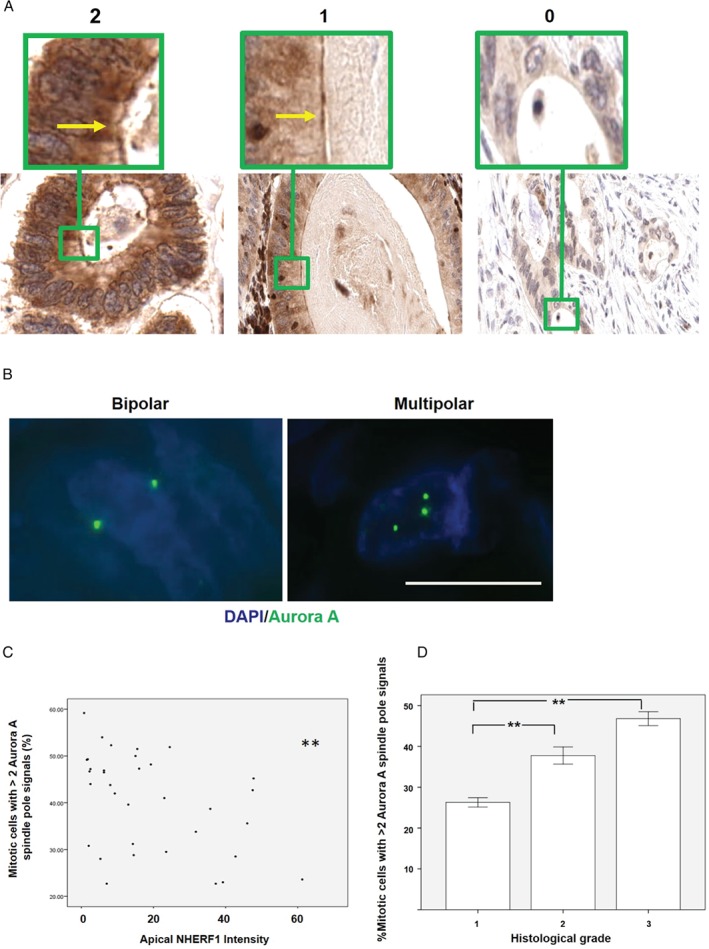
Histological grading provides prognostic stratification of colorectal cancer (CRC) by scoring heterogeneous phenotypes. Features of aggressiveness include aberrant mitotic spindle configurations, chromosomal breakage, and bizarre multicellular morphology, but pathobiology is poorly understood. Protein kinase C zeta (PKCz) controls mitotic spindle dynamics, chromosome segregation, and multicellular patterns, but its role in CRC phenotype evolution remains unclear. Here, we show that PKCz couples genome segregation to multicellular morphology through control of interphase centrosome anchoring. PKCz regulates interdependent processes that control centrosome positioning. Among these, interaction between the cytoskeletal linker protein ezrin and its binding partner NHERF1 promotes the formation of a localized cue for anchoring interphase centrosomes to the cell cortex. Perturbation of these phenomena induced different outcomes in cells with single or extra centrosomes. Defective anchoring of a single centrosome promoted bipolar spindle misorientation, multi-lumen formation, and aberrant epithelial stratification. Collectively, these disturbances induce cribriform multicellular morphology that is typical of some categories of low-grade CRC. By contrast, defective anchoring of extra centrosomes promoted multipolar spindle formation, chromosomal instability (CIN), disruption of glandular morphology, and cell outgrowth across the extracellular matrix interface characteristic of aggressive, high-grade CRC. Because PKCz enhances apical NHERF1 intensity in 3D epithelial cultures, we used an immunohistochemical (IHC) assay of apical NHERF1 intensity as an indirect readout of PKCz activity in translational studies. We show that apical NHERF1 IHC intensity is inversely associated with multipolar spindle frequency and high-grade morphology in formalin-fixed human CRC samples. To conclude, defective PKCz control of interphase centrosome anchoring may underlie distinct categories of mitotic slippage that shape the development of low- or high-grade CRC phenotypes. © 2018 The Authors. The Journal of Pathology published by John Wiley & Sons Ltd on behalf of Pathological Society of Great Britain and Ireland.
Keywords: centrosome; chromosomal instability; colorectal neoplasms; protein kinase C; spindle apparatus.
© 2018 The Authors. The Journal of Pathology published by John Wiley & Sons Ltd on behalf of Pathological Society of Great Britain and Ireland.
Figures
Similar articles
-
Fundamental control of grade-specific colorectal cancer morphology by Src regulation of ezrin-centrosome engagement.J Pathol. 2020 Jul;251(3):310-322. doi: 10.1002/path.5452. Epub 2020 Jun 4. J Pathol. 2020. PMID: 32315081
-
PTEN phosphatase-independent maintenance of glandular morphology in a predictive colorectal cancer model system.Neoplasia. 2013 Nov;15(11):1218-30. doi: 10.1593/neo.121516. Neoplasia. 2013. PMID: 24348097 Free PMC article.
-
NHERF1/EBP50 controls morphogenesis of 3D colonic glands by stabilizing PTEN and ezrin-radixin-moesin proteins at the apical membrane.Neoplasia. 2014 Apr;16(4):365-74.e1-2. doi: 10.1016/j.neo.2014.04.004. Neoplasia. 2014. PMID: 24862762 Free PMC article.
-
Centrosomes, chromosome instability (CIN) and aneuploidy.Curr Opin Cell Biol. 2012 Dec;24(6):809-15. doi: 10.1016/j.ceb.2012.10.006. Epub 2012 Nov 3. Curr Opin Cell Biol. 2012. PMID: 23127609 Free PMC article. Review.
-
The role of mitotic kinases in coupling the centrosome cycle with the assembly of the mitotic spindle.J Cell Sci. 2014 Oct 1;127(Pt 19):4111-22. doi: 10.1242/jcs.151753. Epub 2014 Aug 15. J Cell Sci. 2014. PMID: 25128564 Review.
Cited by
-
Methylation-mediated silencing of protein kinase C zeta induces apoptosis avoidance through ATM/CHK2 inactivation in dedifferentiated chondrosarcoma.Br J Cancer. 2022 May;126(9):1289-1300. doi: 10.1038/s41416-021-01695-1. Epub 2022 Jan 11. Br J Cancer. 2022. PMID: 35017658 Free PMC article.
-
Mechanistic Insights into Colorectal Cancer Phenomics from Fundamental and Organotypic Model Studies.Am J Pathol. 2018 Sep;188(9):1936-1948. doi: 10.1016/j.ajpath.2018.05.021. Epub 2018 Jul 18. Am J Pathol. 2018. PMID: 30028958 Free PMC article. Review.
References
-
- Arnold M, Sierra MS, Laversanne M, et al Global patterns and trends in colorectal cancer incidence and mortality. Gut 2017; 66 : 683–691. - PubMed
-
- Nicholl ID, Dunlop MG. Molecular markers of prognosis in colorectal cancer. J Natl Cancer Inst 1999; 91 : 1267–1269. - PubMed
-
- Jass JR, Atkin WS, Cuzick J, et al The grading of rectal cancer: historical perspectives and a multivariate analysis of 447 cases. Histopathology 1986; 10 : 437–459. - PubMed
-
- Ueno H, Hase K, Hashiguchi Y, et al Site‐specific tumor grading system in colorectal cancer: multicenter pathologic review of the value of quantifying poorly differentiated clusters. Am J Surg Pathol 2014; 38 : 197–204. - PubMed
-
- Walther A, Houlston R, Tomlinson I. Association between chromosomal instability and prognosis in colorectal cancer: a meta‐analysis. Gut 2008; 57 : 941–950. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
