

Morphogenetic defects underlie Superior Coloboma, a newly identified closure disorder of the dorsal eye
- PMID: 29522511
- PMCID: PMC5862500
- DOI: 10.1371/journal.pgen.1007246
Morphogenetic defects underlie Superior Coloboma, a newly identified closure disorder of the dorsal eye
Abstract
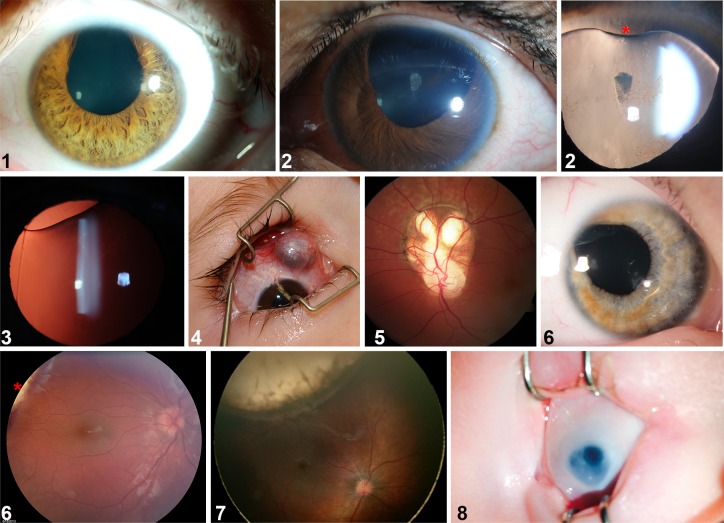
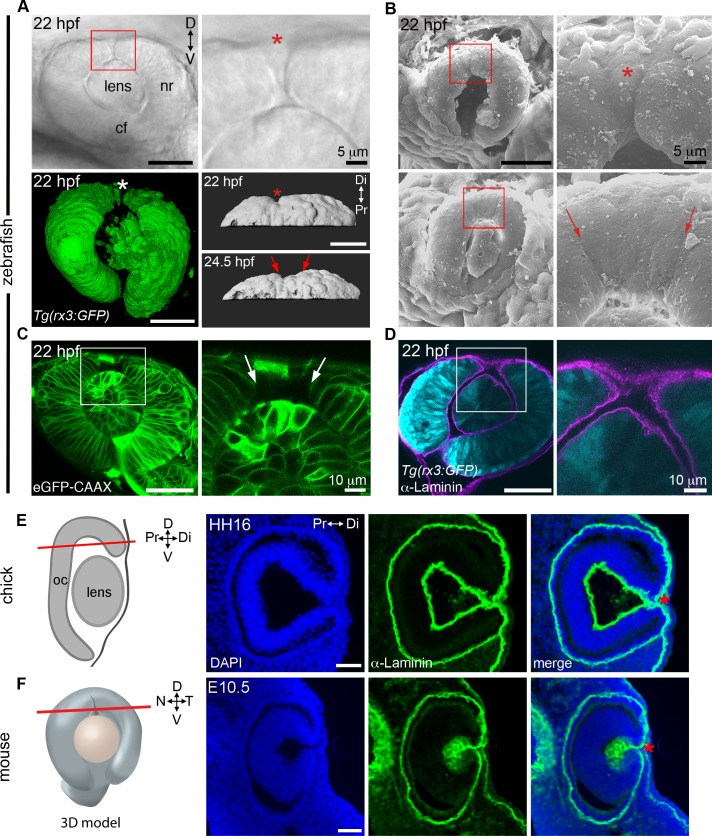
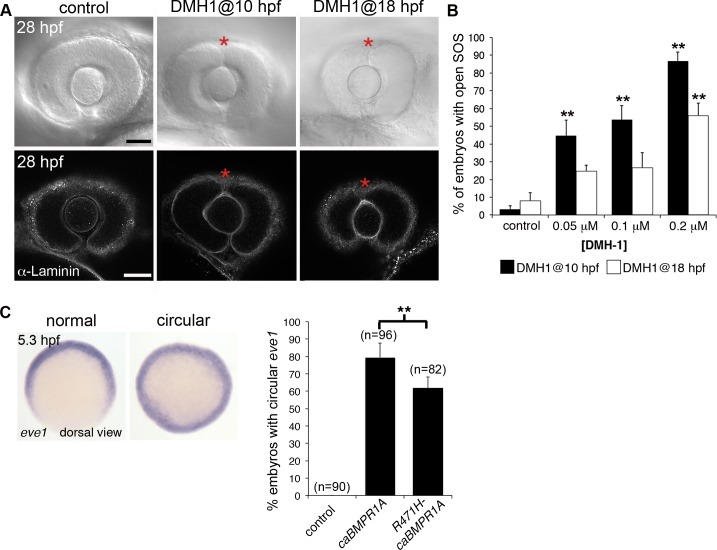
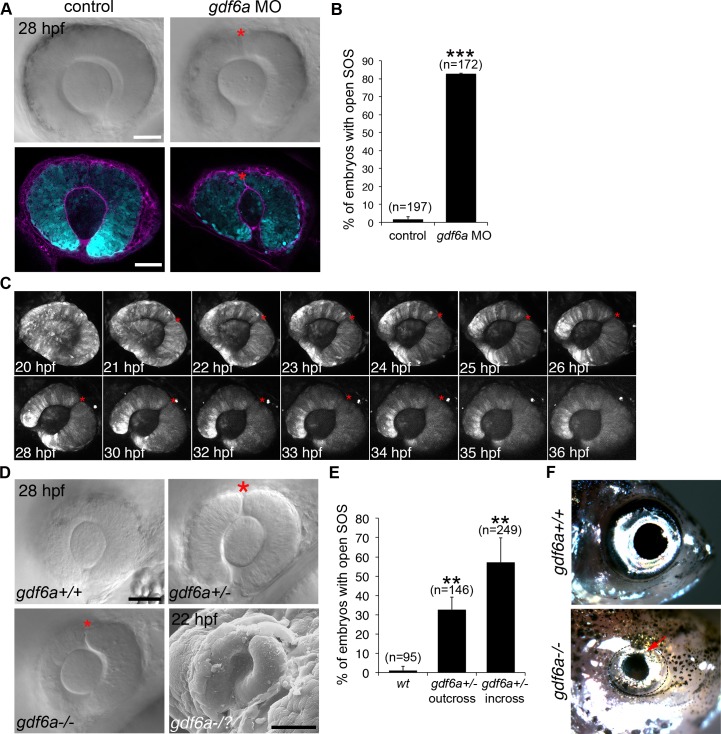
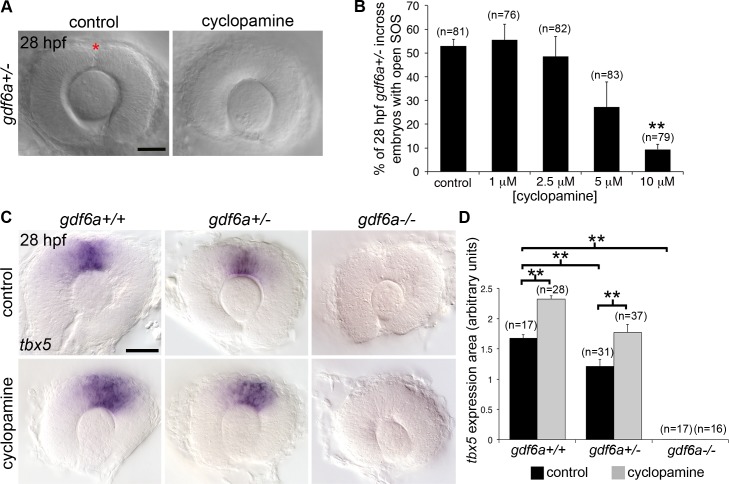
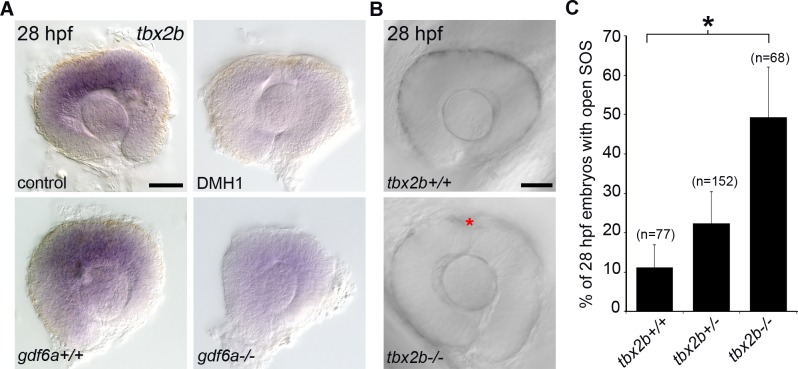
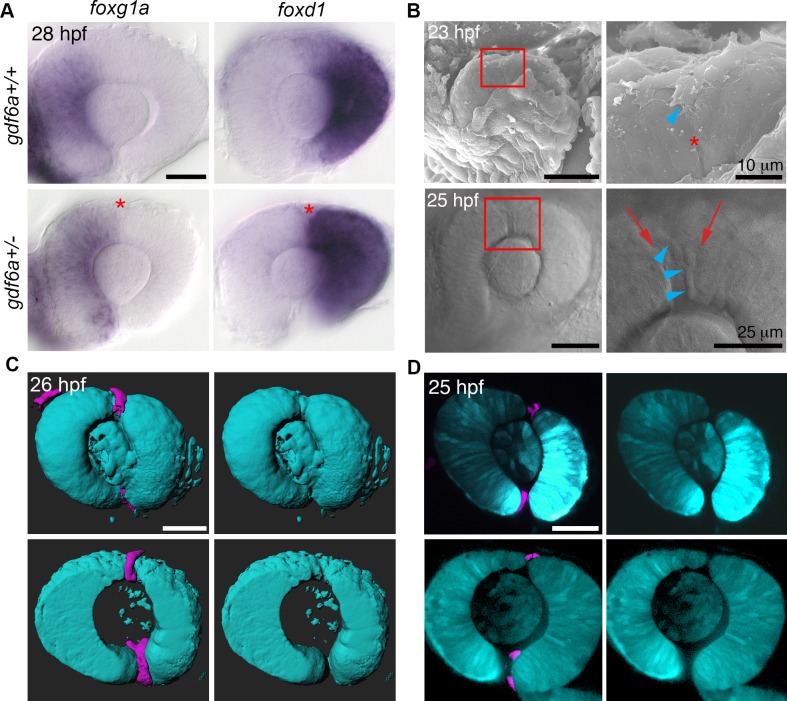
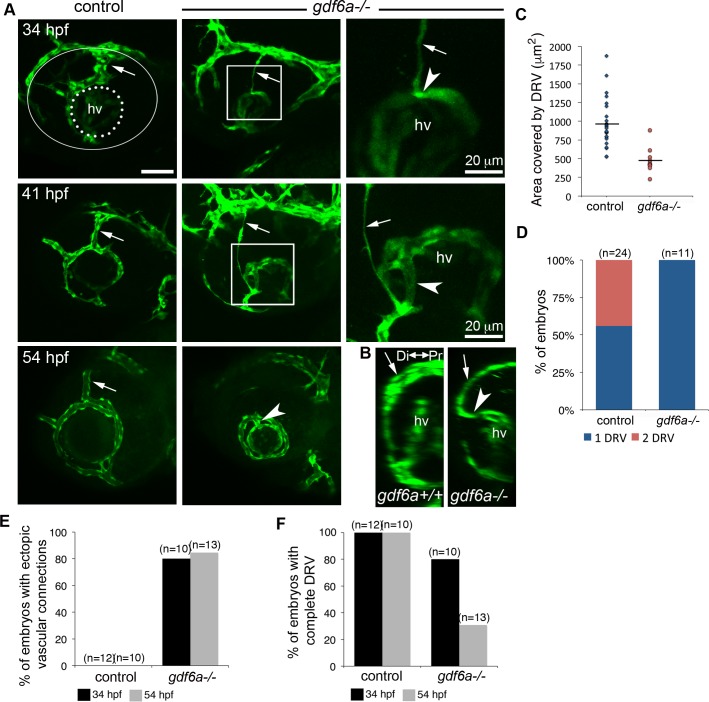
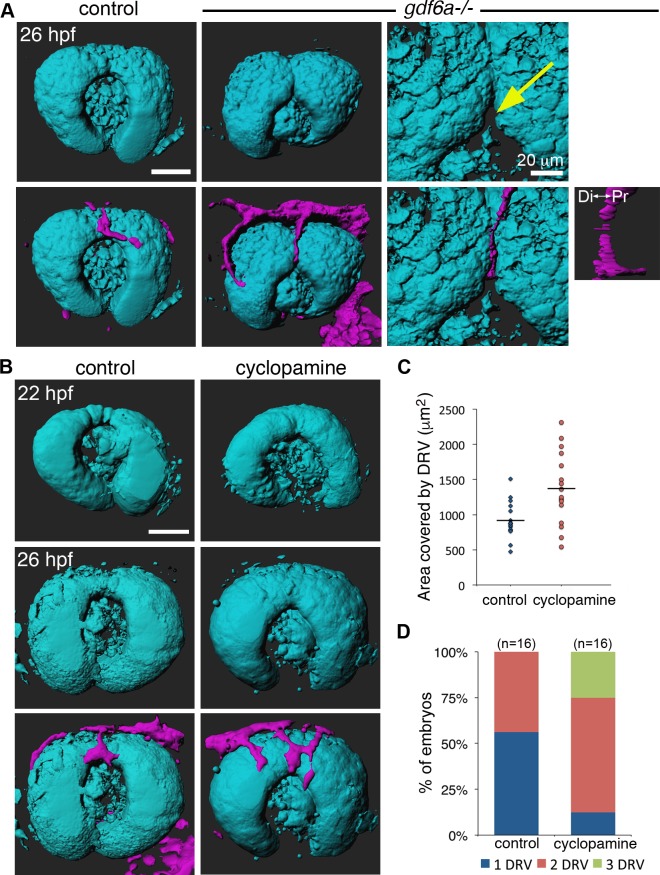
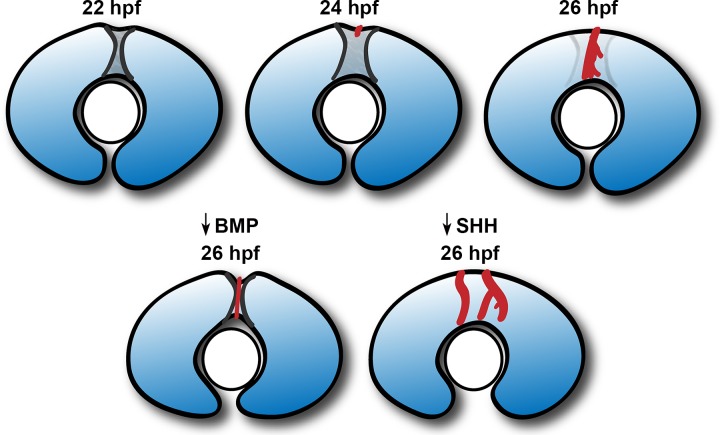
The eye primordium arises as a lateral outgrowth of the forebrain, with a transient fissure on the inferior side of the optic cup providing an entry point for developing blood vessels. Incomplete closure of the inferior ocular fissure results in coloboma, a disease characterized by gaps in the inferior eye and recognized as a significant cause of pediatric blindness. Here, we identify eight patients with defects in tissues of the superior eye, a congenital disorder that we term superior coloboma. The embryonic origin of superior coloboma could not be explained by conventional models of eye development, leading us to reanalyze morphogenesis of the dorsal eye. Our studies revealed the presence of the superior ocular sulcus (SOS), a transient division of the dorsal eye conserved across fish, chick, and mouse. Exome sequencing of superior coloboma patients identified rare variants in a Bone Morphogenetic Protein (Bmp) receptor (BMPR1A) and T-box transcription factor (TBX2). Consistent with this, we find sulcus closure defects in zebrafish lacking Bmp signaling or Tbx2b. In addition, loss of dorsal ocular Bmp is rescued by concomitant suppression of the ventral-specific Hedgehog pathway, arguing that sulcus closure is dependent on dorsal-ventral eye patterning cues. The superior ocular sulcus acts as a conduit for blood vessels, with altered sulcus closure resulting in inappropriate connections between the hyaloid and superficial vascular systems. Together, our findings explain the existence of superior coloboma, a congenital ocular anomaly resulting from aberrant morphogenesis of a developmental structure.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Morphogenesis and axis specification occur in parallel during optic cup and optic fissure formation, differentially modulated by BMP and Wnt.Open Biol. 2019 Feb 28;9(2):180179. doi: 10.1098/rsob.180179. Open Biol. 2019. PMID: 30958096 Free PMC article.
-
Sox4 regulates choroid fissure closure by limiting Hedgehog signaling during ocular morphogenesis.Dev Biol. 2015 Mar 1;399(1):139-153. doi: 10.1016/j.ydbio.2014.12.026. Epub 2014 Dec 31. Dev Biol. 2015. PMID: 25557621 Free PMC article.
-
Abnormal vasculature interferes with optic fissure closure in lmo2 mutant zebrafish embryos.Dev Biol. 2012 Sep 15;369(2):191-8. doi: 10.1016/j.ydbio.2012.06.029. Epub 2012 Jul 20. Dev Biol. 2012. PMID: 22819672
-
Genes and pathways in optic fissure closure.Semin Cell Dev Biol. 2019 Jul;91:55-65. doi: 10.1016/j.semcdb.2017.10.010. Epub 2017 Dec 6. Semin Cell Dev Biol. 2019. PMID: 29198497 Review.
-
Ocular coloboma: a reassessment in the age of molecular neuroscience.J Med Genet. 2004 Dec;41(12):881-91. doi: 10.1136/jmg.2004.025494. J Med Genet. 2004. PMID: 15591273 Free PMC article. Review.
Cited by
-
A rare case of temporal atypical retinochoroidal coloboma associated with posterior embryotoxon.Indian J Ophthalmol. 2020 Jul;68(7):1445-1446. doi: 10.4103/ijo.IJO_2213_19. Indian J Ophthalmol. 2020. PMID: 32587187 Free PMC article. No abstract available.
-
In Vivo Analysis of Optic Fissure Fusion in Zebrafish: Pioneer Cells, Basal Lamina, Hyaloid Vessels, and How Fissure Fusion is Affected by BMP.Int J Mol Sci. 2020 Apr 16;21(8):2760. doi: 10.3390/ijms21082760. Int J Mol Sci. 2020. PMID: 32316164 Free PMC article.
-
Cercosporamide inhibits bone morphogenetic protein receptor type I kinase activity in zebrafish.Dis Model Mech. 2020 Sep 24;13(9):dmm045971. doi: 10.1242/dmm.045971. Dis Model Mech. 2020. PMID: 32820031 Free PMC article.
-
Novel insights into chorioretinal and juxtapapillary colobomas by optical coherence tomography.Vet Ophthalmol. 2022 May;25 Suppl 1(Suppl 1):136-143. doi: 10.1111/vop.12970. Epub 2022 Jan 29. Vet Ophthalmol. 2022. PMID: 35092136 Free PMC article.
-
Morphogenesis and axis specification occur in parallel during optic cup and optic fissure formation, differentially modulated by BMP and Wnt.Open Biol. 2019 Feb 28;9(2):180179. doi: 10.1098/rsob.180179. Open Biol. 2019. PMID: 30958096 Free PMC article.
References
-
- Bazin-Lopez N, Valdivia LE, Wilson SW, Gestri G. Watching eyes take shape. Curr Opin Genet Dev. 2015;32: 73–79. doi: 10.1016/j.gde.2015.02.004 - DOI - PMC - PubMed
-
- Fuhrmann S. Eye morphogenesis and patterning of the optic vesicle. Curr Top Dev Biol. 2010;93: 61–84. doi: 10.1016/B978-0-12-385044-7.00003-5 - DOI - PMC - PubMed
-
- Chang L, Blain D, Bertuzzi S, Brooks BP. Uveal coloboma: clinical and basic science update. Curr Opin Ophthalmol. 2006;17: 447–470. doi: 10.1097/01.icu.0000243020.82380.f6 - DOI - PubMed
-
- Onwochei BC, Simon JW, Bateman JB, Couture KC, Mir E. Ocular colobomata. Surv Ophthalmol. 2000;45: 175–194. - PubMed
-
- Williamson KA, FitzPatrick DR. The genetic architecture of microphthalmia, anophthalmia and coloboma. Eur J Med Genet. 2014;57: 369–380. doi: 10.1016/j.ejmg.2014.05.002 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
