

Lung macrophage scavenger receptor SR-A6 (MARCO) is an adenovirus type-specific virus entry receptor
- PMID: 29522575
- PMCID: PMC5862501
- DOI: 10.1371/journal.ppat.1006914
Lung macrophage scavenger receptor SR-A6 (MARCO) is an adenovirus type-specific virus entry receptor
Abstract
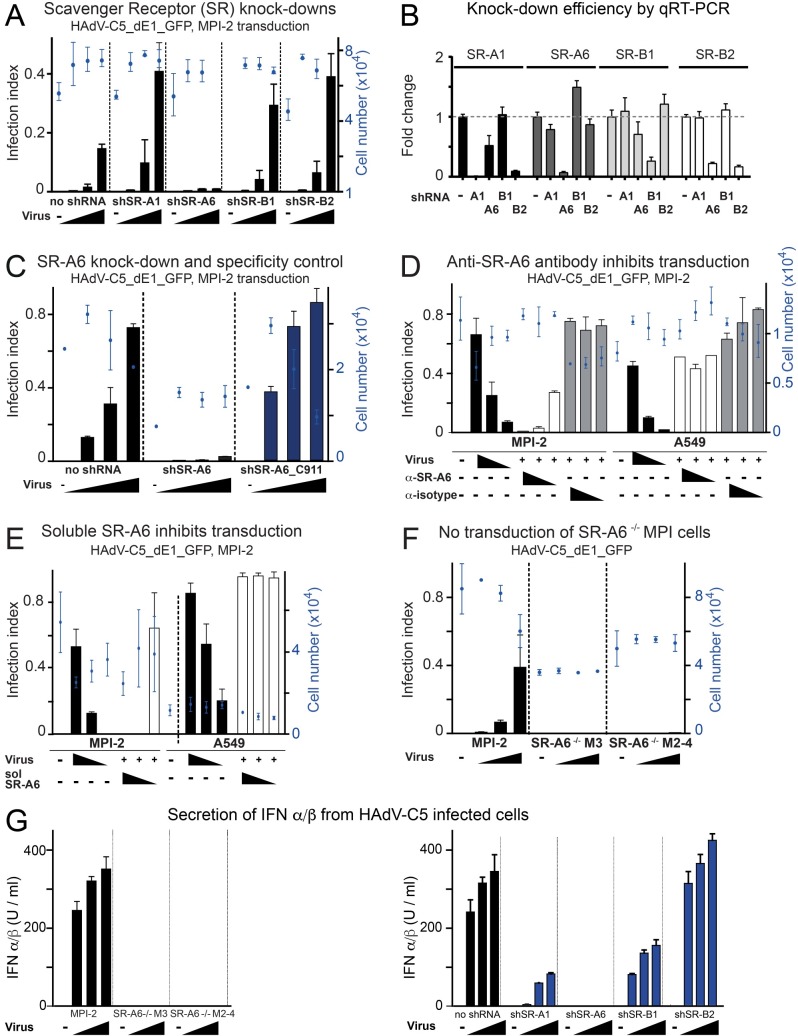
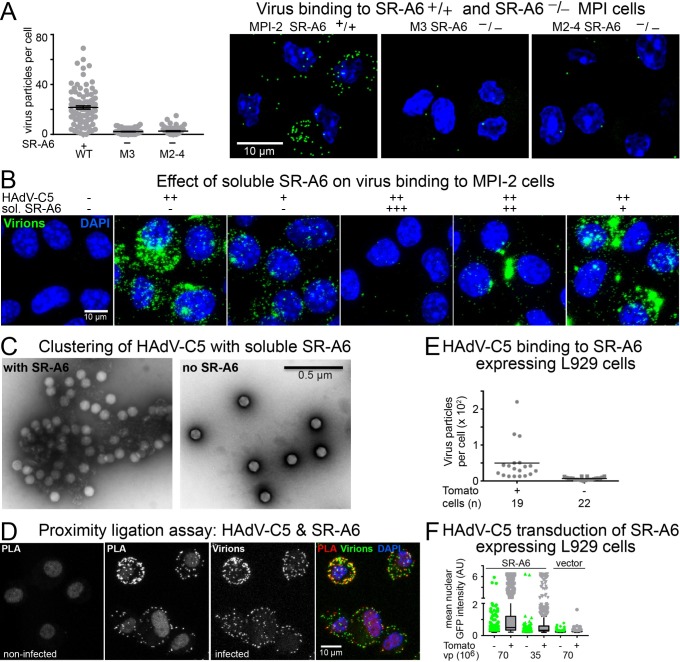
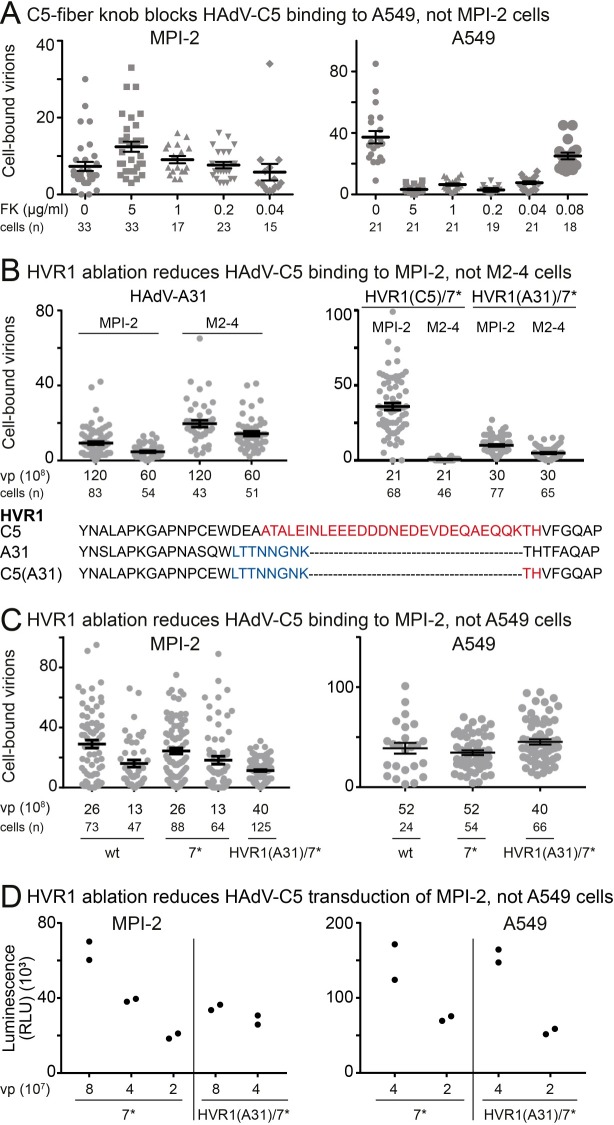
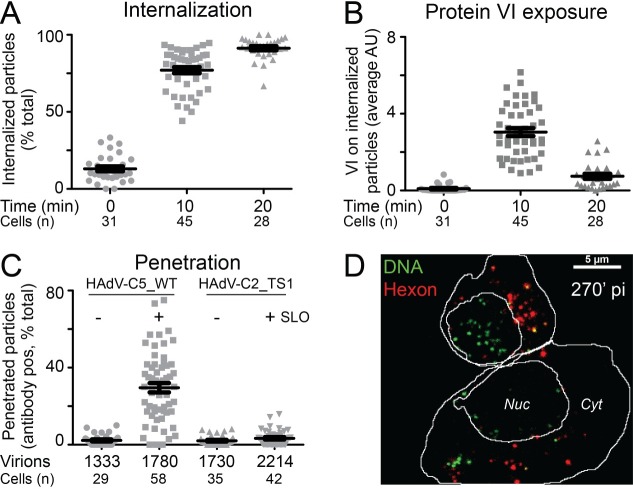
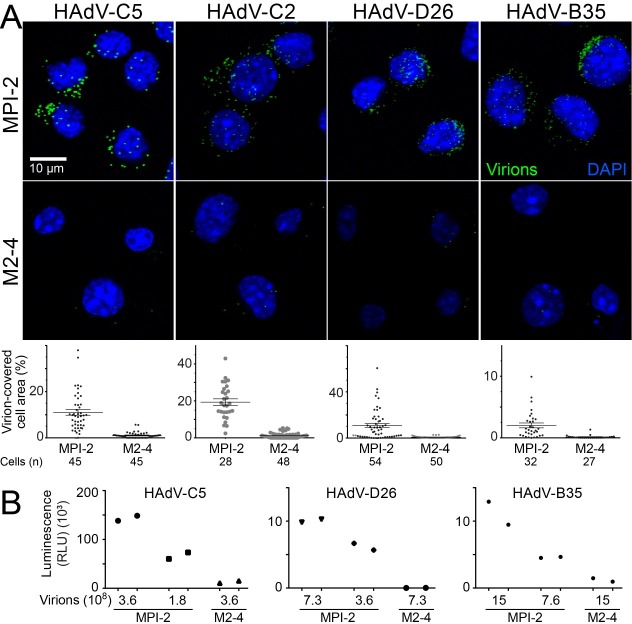
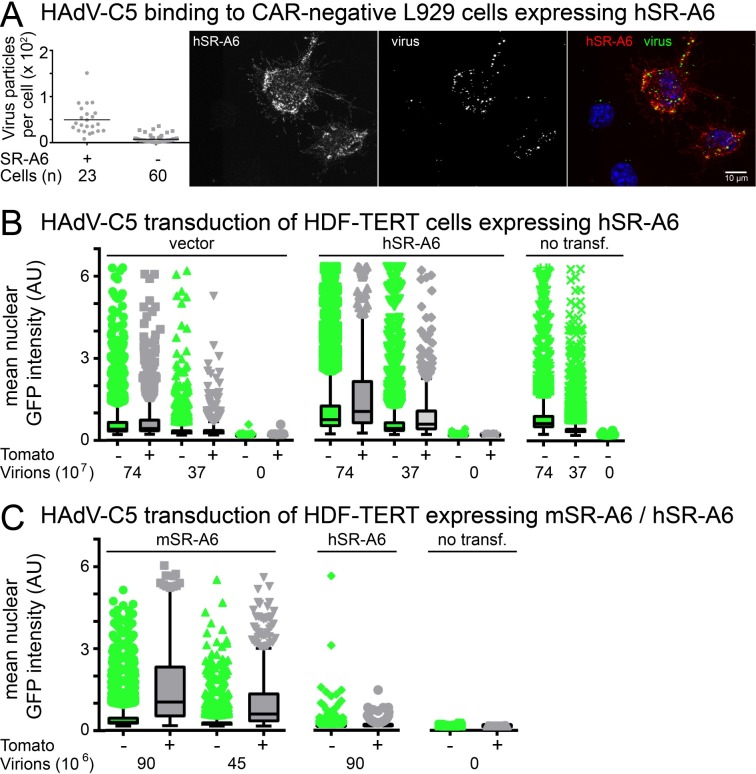
Macrophages are a diverse group of phagocytic cells acting in host protection against stress, injury, and pathogens. Here, we show that the scavenger receptor SR-A6 is an entry receptor for human adenoviruses in murine alveolar macrophage-like MPI cells, and important for production of type I interferon. Scavenger receptors contribute to the clearance of endogenous proteins, lipoproteins and pathogens. Knockout of SR-A6 in MPI cells, anti-SR-A6 antibody or the soluble extracellular SR-A6 domain reduced adenovirus type-C5 (HAdV-C5) binding and transduction. Expression of murine SR-A6, and to a lower extent human SR-A6 boosted virion binding to human cells and transduction. Virion clustering by soluble SR-A6 and proximity localization with SR-A6 on MPI cells suggested direct adenovirus interaction with SR-A6. Deletion of the negatively charged hypervariable region 1 (HVR1) of hexon reduced HAdV-C5 binding and transduction, implying that the viral ligand for SR-A6 is hexon. SR-A6 facilitated macrophage entry of HAdV-B35 and HAdV-D26, two important vectors for transduction of hematopoietic cells and human vaccination. The study highlights the importance of scavenger receptors in innate immunity against human viruses.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Shi C, Pamer EG. Monocyte recruitment during infection and inflammation. Nat Rev Immunol. 2011;11(11):762–74. doi: 10.1038/nri3070 ; PubMed Central PMCID: PMCPMC3947780. - DOI - PMC - PubMed
-
- Davies LC, Jenkins SJ, Allen JE, Taylor PR. Tissue-resident macrophages. Nat Immunol. 2013;14(10):986–95. doi: 10.1038/ni.2705 ; PubMed Central PMCID: PMCPMC4045180. - DOI - PMC - PubMed
-
- Price JV, Vance RE. The Macrophage Paradox. Immunity. 2014;41(5):685–93. Epub 2014/12/18. doi: 10.1016/j.immuni.2014.10.015 . - DOI - PubMed
-
- Ginhoux F, Schultze JL, Murray PJ, Ochando J, Biswas SK. New insights into the multidimensional concept of macrophage ontogeny, activation and function. Nat Immunol. 2016;17(1):34–40. doi: 10.1038/ni.3324 . - DOI - PubMed
-
- Verschoor CP, Puchta A, Bowdish DM. The macrophage. Methods Mol Biol. 2012;844:139–56. Epub 2012/01/21. doi: 10.1007/978-1-61779-527-5_10 . - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
