

Mbf1 ensures Polycomb silencing by protecting E(z) mRNA from degradation by Pacman
- PMID: 29523653
- PMCID: PMC5868998
- DOI: 10.1242/dev.162461
Mbf1 ensures Polycomb silencing by protecting E(z) mRNA from degradation by Pacman
Abstract
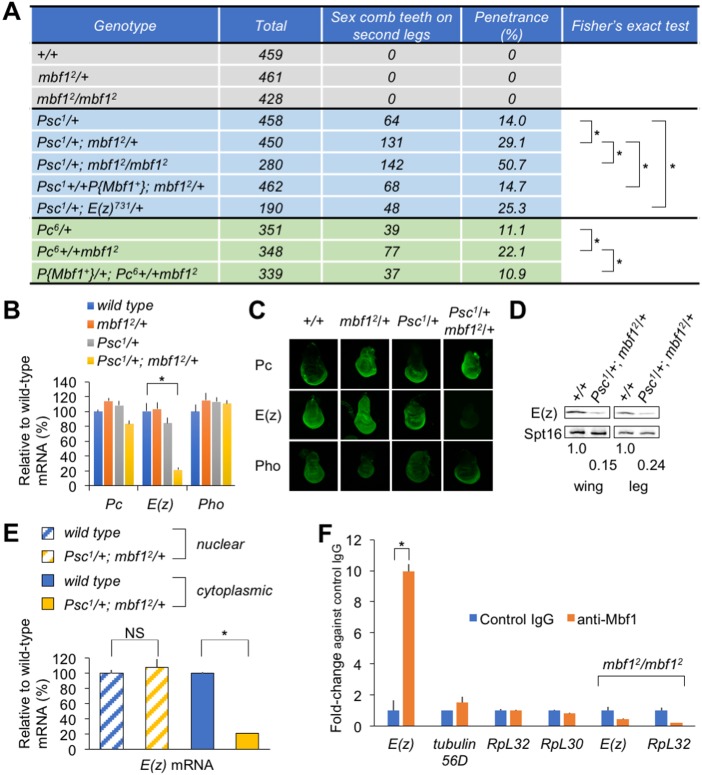
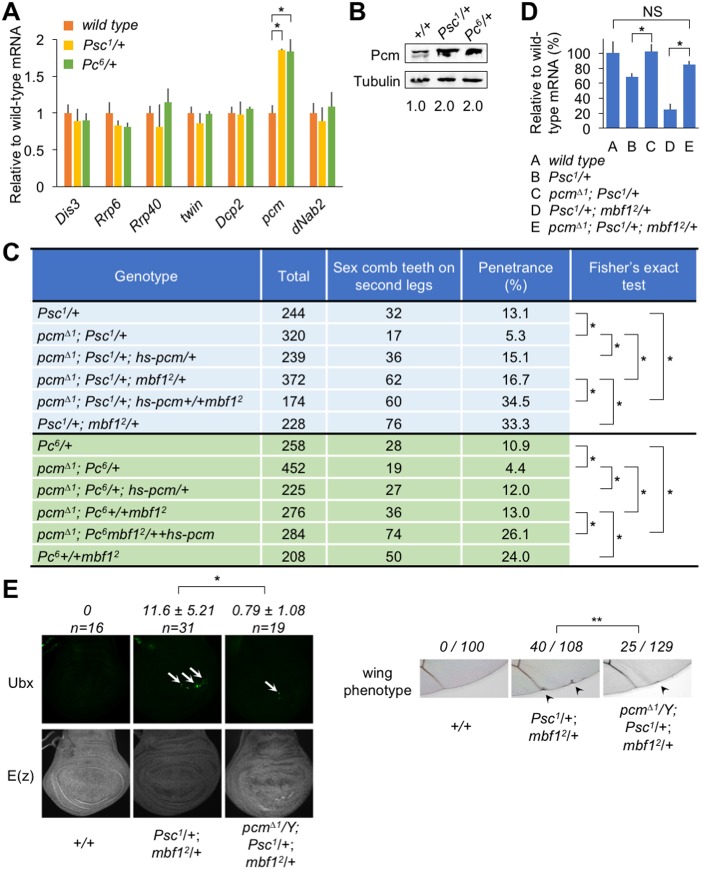
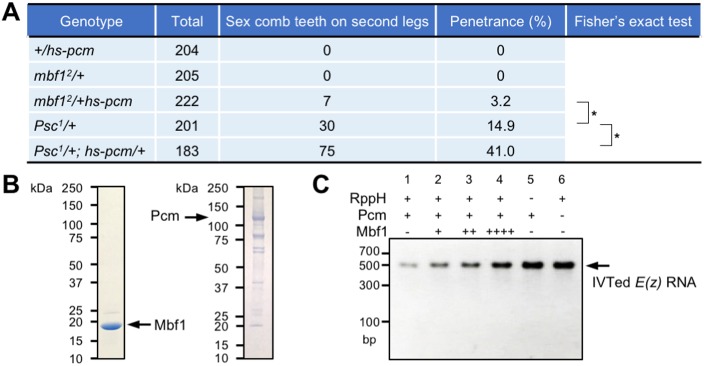
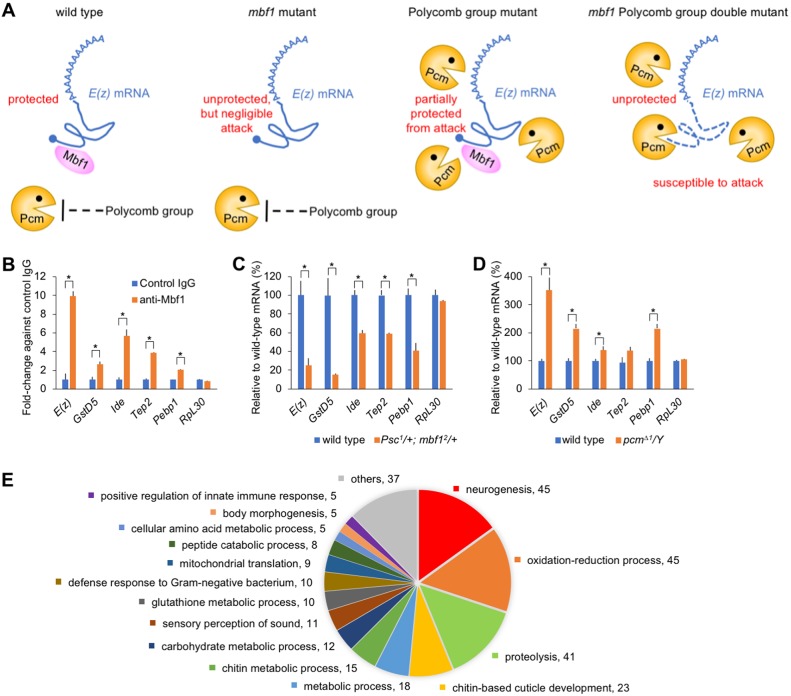
Under stress conditions, the coactivator Multiprotein bridging factor 1 (Mbf1) translocates from the cytoplasm into the nucleus to induce stress-response genes. However, its role in the cytoplasm, where it is mainly located, has remained elusive. Here, we show that Drosophila Mbf1 associates with E(z) mRNA and protects it from degradation by the exoribonuclease Pacman (Pcm), thereby ensuring Polycomb silencing. In genetic studies, loss of mbf1 function enhanced a Polycomb phenotype in Polycomb group mutants, and was accompanied by a significant reduction in E(z) mRNA expression. Furthermore, a pcm mutation suppressed the Polycomb phenotype and restored the expression level of E(z) mRNA, while pcm overexpression exhibited the Polycomb phenotype in the mbf1 mutant but not in the wild-type background. In vitro, Mbf1 protected E(z) RNA from Pcm activity. Our results suggest that Mbf1 buffers fluctuations in Pcm activity to maintain an E(z) mRNA expression level sufficient for Polycomb silencing.
Keywords: Drosophila melanogaster; E(z); Enhancer of zeste; Exoribonuclease; Mbf1; Pcm; Polycomb silencing.
© 2018. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
Similar articles
-
Distinct functions of polycomb group proteins in regulating Ci transcription in developing Drosophila.J Mol Cell Biol. 2018 Oct 1;10(5):475-478. doi: 10.1093/jmcb/mjy009. J Mol Cell Biol. 2018. PMID: 29924323 No abstract available.
-
Drosophila MBF1 is a co-activator for Tracheae Defective and contributes to the formation of tracheal and nervous systems.Development. 2003 Feb;130(4):719-28. doi: 10.1242/dev.00297. Development. 2003. PMID: 12506002
-
SIR2 is required for polycomb silencing and is associated with an E(Z) histone methyltransferase complex.Curr Biol. 2004 Oct 26;14(20):1812-21. doi: 10.1016/j.cub.2004.09.060. Curr Biol. 2004. PMID: 15498488
-
[Coactivator MBF1 governs oxidative stress response].Tanpakushitsu Kakusan Koso. 2005 Feb;50(2):136-40. Tanpakushitsu Kakusan Koso. 2005. PMID: 15704460 Review. Japanese. No abstract available.
-
The functions of E(Z)/EZH2-mediated methylation of lysine 27 in histone H3.Curr Opin Genet Dev. 2004 Apr;14(2):155-64. doi: 10.1016/j.gde.2004.02.001. Curr Opin Genet Dev. 2004. PMID: 15196462 Review.
Cited by
-
Unbiased shRNA screening, using a combination of FACS and high-throughput sequencing, enables identification of novel modifiers of Polycomb silencing.Sci Rep. 2018 Aug 14;8(1):12128. doi: 10.1038/s41598-018-30649-6. Sci Rep. 2018. PMID: 30108332 Free PMC article.
-
Sex-dimorphic tumor growth is regulated by tumor microenvironmental and systemic signals.Sci Adv. 2024 Dec 6;10(49):eads4229. doi: 10.1126/sciadv.ads4229. Epub 2024 Dec 6. Sci Adv. 2024. PMID: 39642218 Free PMC article.
-
Discovering the RNA-Binding Proteome of Plant Leaves with an Improved RNA Interactome Capture Method.Biomolecules. 2020 Apr 24;10(4):661. doi: 10.3390/biom10040661. Biomolecules. 2020. PMID: 32344669 Free PMC article.
References
-
- Arce D. P., Godoy A. V., Tsuda K., Yamazaki K., Valle E. M., Iglesias M. J., Di Mauro M. F. and Casalongué C. A. (2010). The analysis of an Arabidopsis triple knock-down mutant reveals functions for MBF1 genes under oxidative stress conditions. J. Plant Physiol. 167, 194-200. 10.1016/j.jplph.2009.09.003 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
