

A Restricted Role for FcγR in the Regulation of Adaptive Immunity
- PMID: 29523656
- PMCID: PMC5896742
- DOI: 10.4049/jimmunol.1700429
A Restricted Role for FcγR in the Regulation of Adaptive Immunity
Abstract
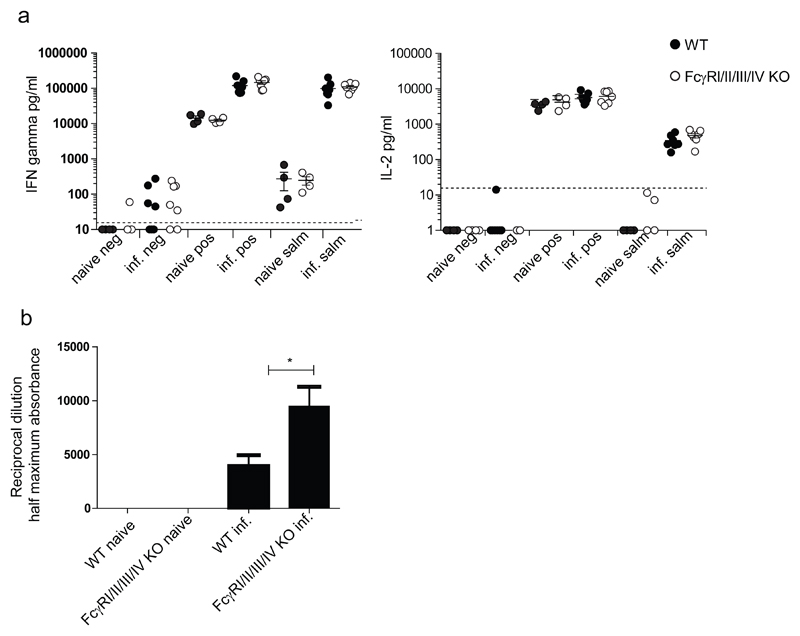
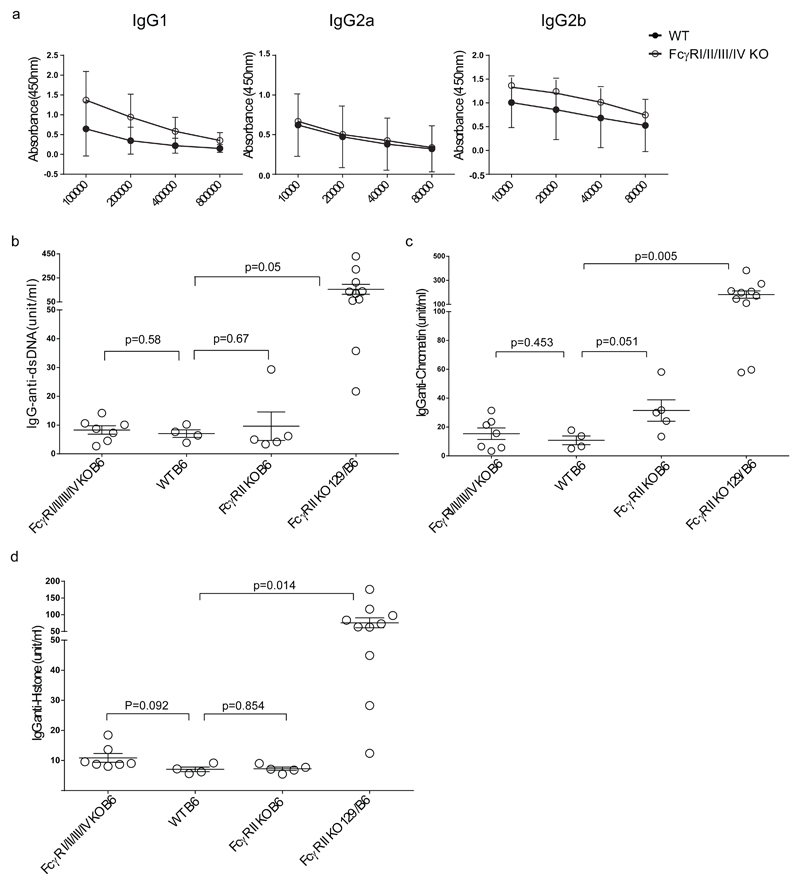
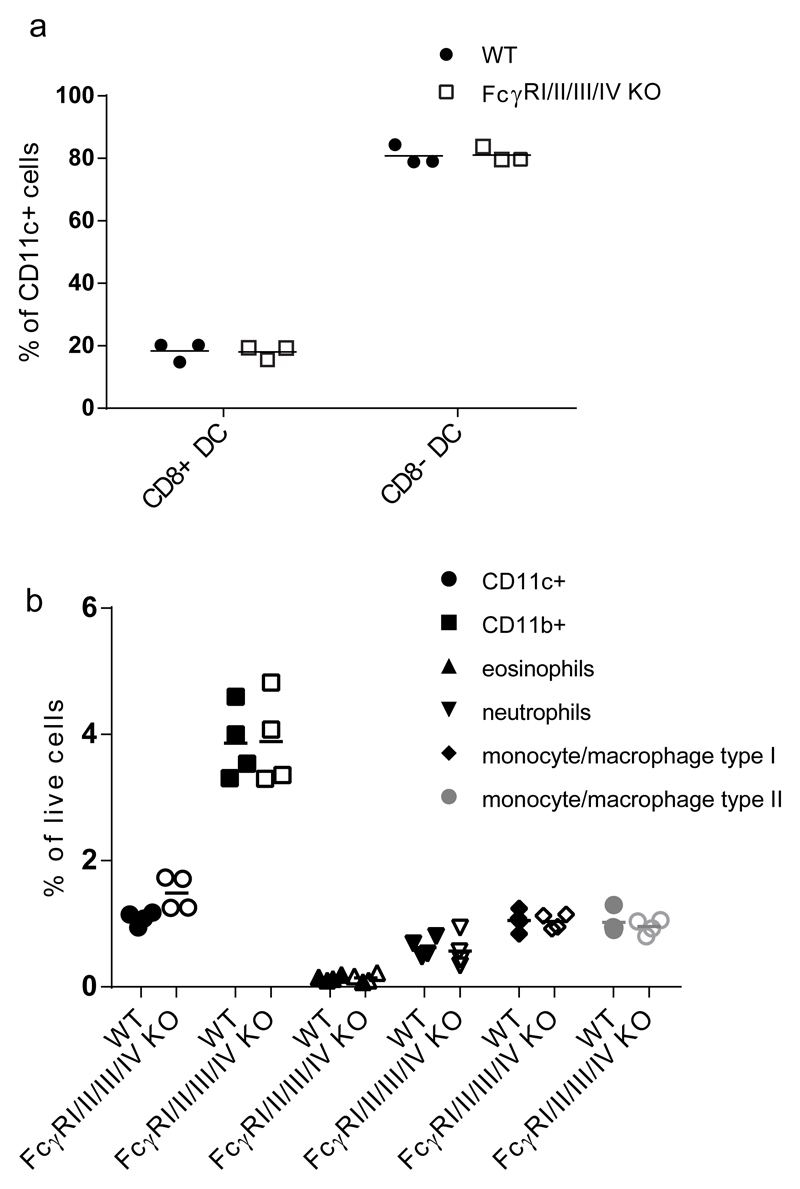
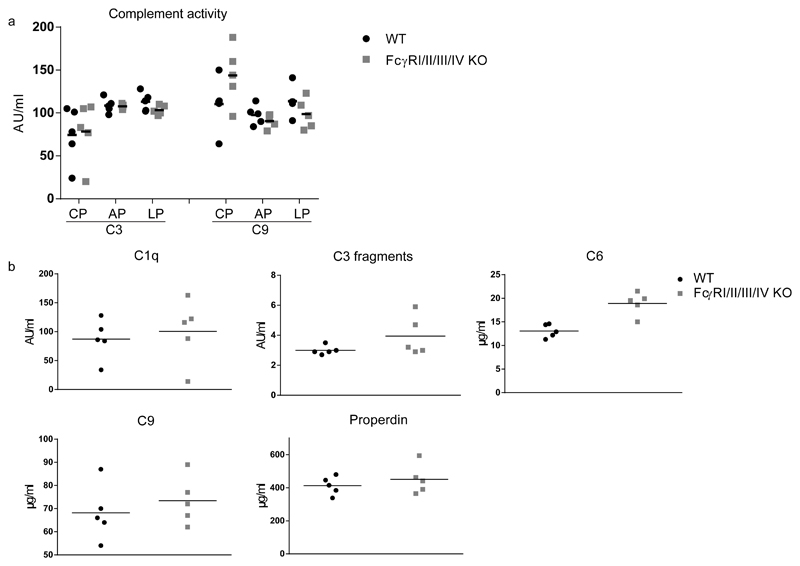
By their interaction with IgG immune complexes, FcγR and complement link innate and adaptive immunity, showing functional redundancy. In complement-deficient mice, IgG downstream effector functions are often impaired, as well as adaptive immunity. Based on a variety of model systems using FcγR-knockout mice, it has been concluded that FcγRs are also key regulators of innate and adaptive immunity; however, several of the model systems underpinning these conclusions suffer from flawed experimental design. To address this issue, we generated a novel mouse model deficient for all FcγRs (FcγRI/II/III/IV-/- mice). These mice displayed normal development and lymphoid and myeloid ontogeny. Although IgG effector pathways were impaired, adaptive immune responses to a variety of challenges, including bacterial infection and IgG immune complexes, were not. Like FcγRIIb-deficient mice, FcγRI/II/III/IV-/- mice developed higher Ab titers but no autoantibodies. These observations indicate a redundant role for activating FcγRs in the modulation of the adaptive immune response in vivo. We conclude that FcγRs are downstream IgG effector molecules with a restricted role in the ontogeny and maintenance of the immune system, as well as the regulation of adaptive immunity.
Copyright © 2018 by The American Association of Immunologists, Inc.
Figures
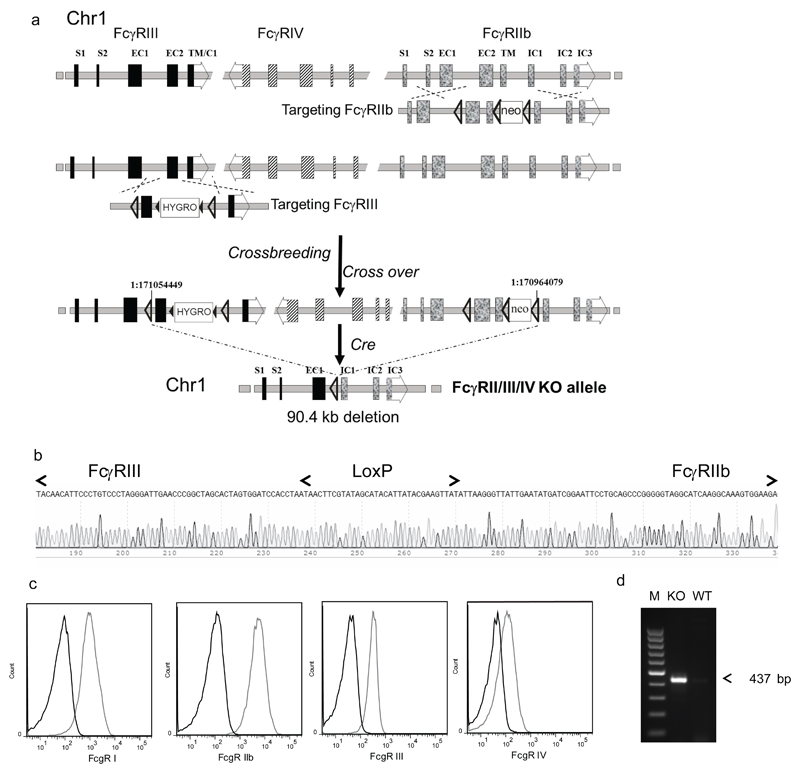
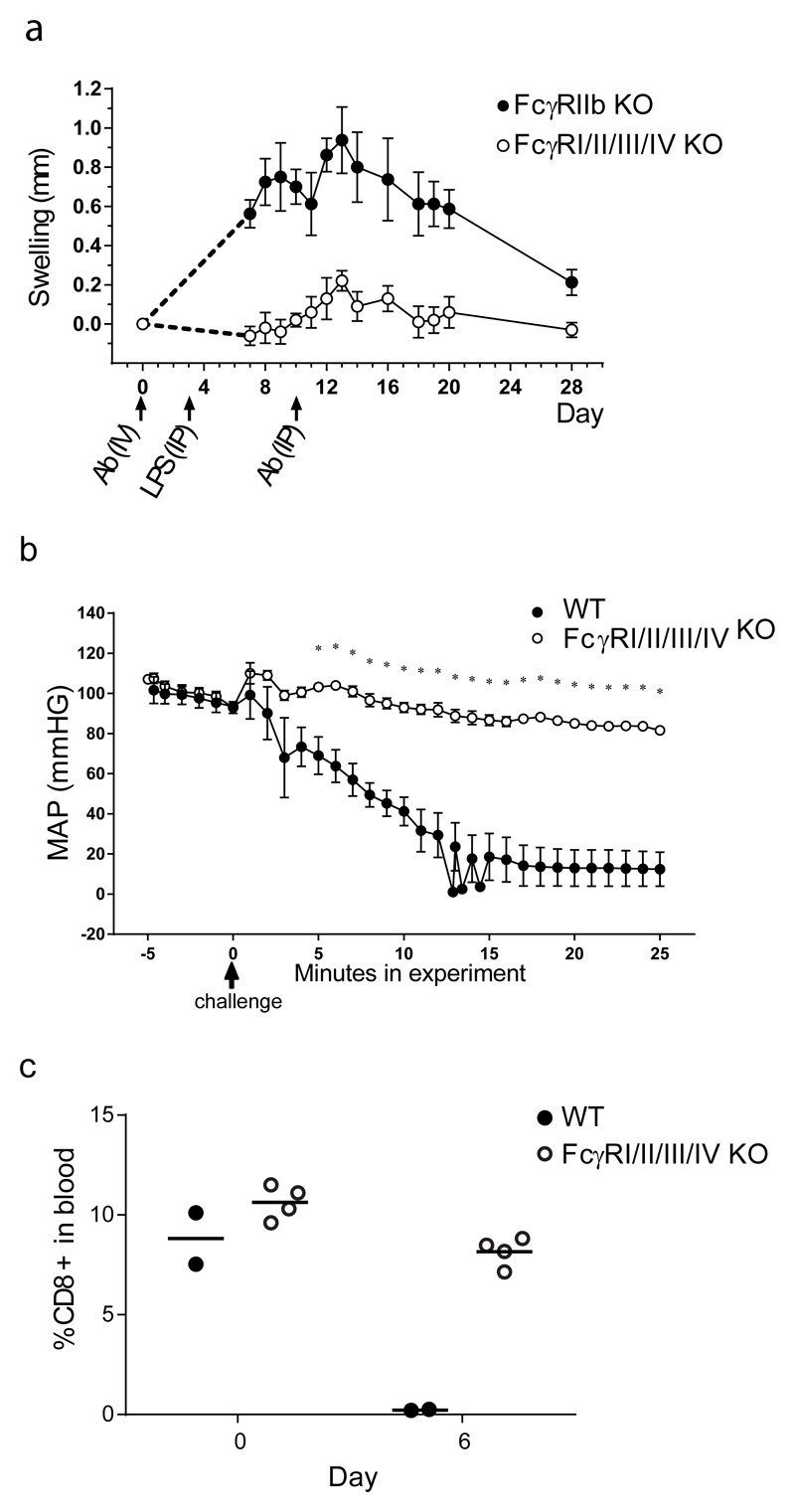
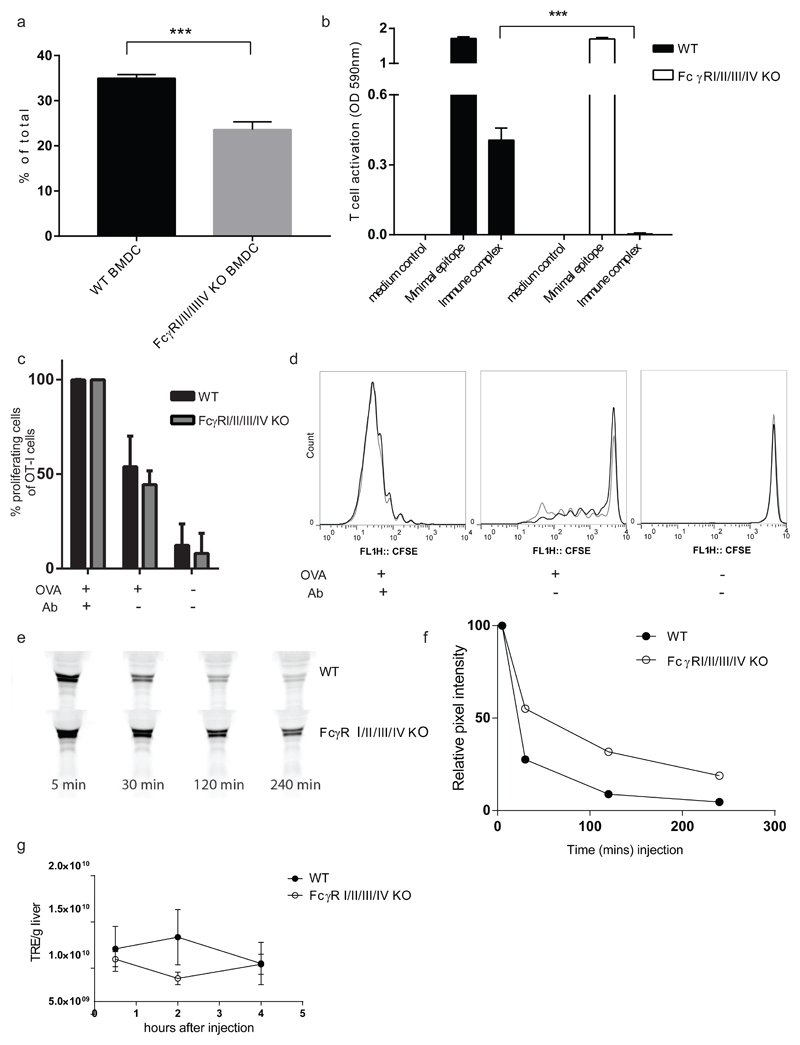
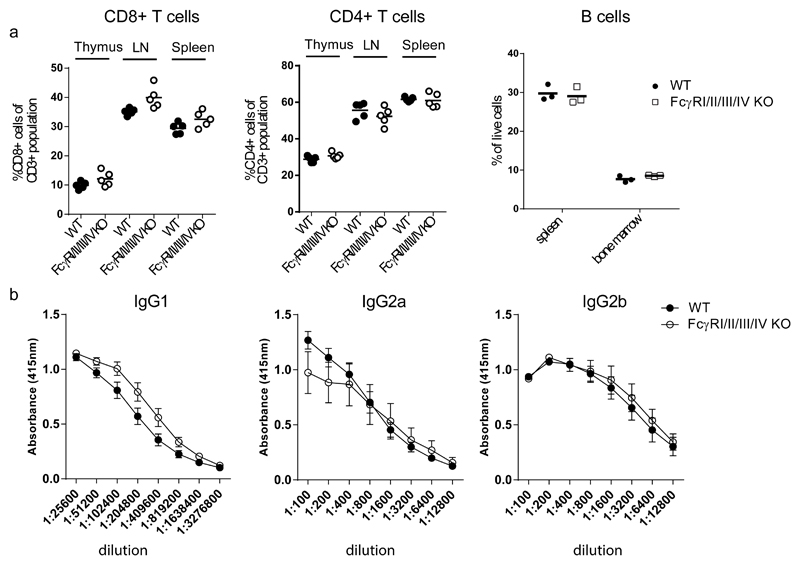
References
-
- Takai T, Li M, Sylvestre D, Clynes R, Ravetch JV. FcR gamma chain deletion results in pleiotrophic effector cell defects. Cell. 1994;76:519–529. - PubMed
-
- Bruhns P, Jonsson F. Mouse and human FcR effector functions. Immunol Rev. 2015;268:25–51. - PubMed
-
- Fearon DT, Carroll MC. Regulation of B lymphocyte responses to foreign and self-antigens by the CD19/CD21 complex. Annu Rev Immunol. 2000;18:393–422. - PubMed
-
- Ahearn JM, Fischer MB, Croix D, Goerg S, Ma M, Xia J, Zhou X, Howard RG, Rothstein TL, Carroll MC. Disruption of the Cr2 locus results in a reduction in B-1a cells and in an impaired B cell response to T-dependent antigen. Immunity. 1996;4:251–262. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
