

Redundant regulation of localization and protein stability of DmPar3
- PMID: 29523893
- PMCID: PMC11105499
- DOI: 10.1007/s00018-018-2792-1
Redundant regulation of localization and protein stability of DmPar3
Abstract
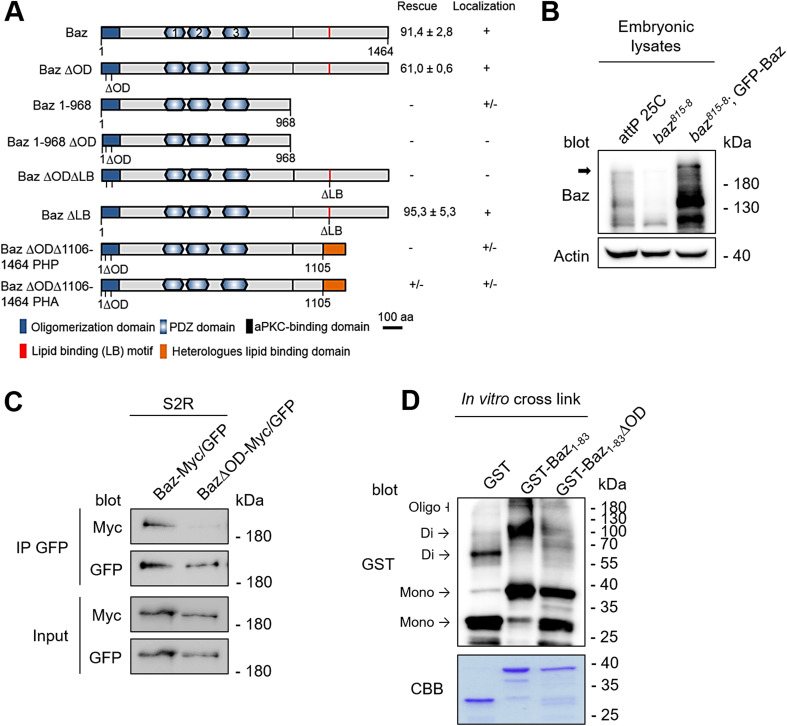
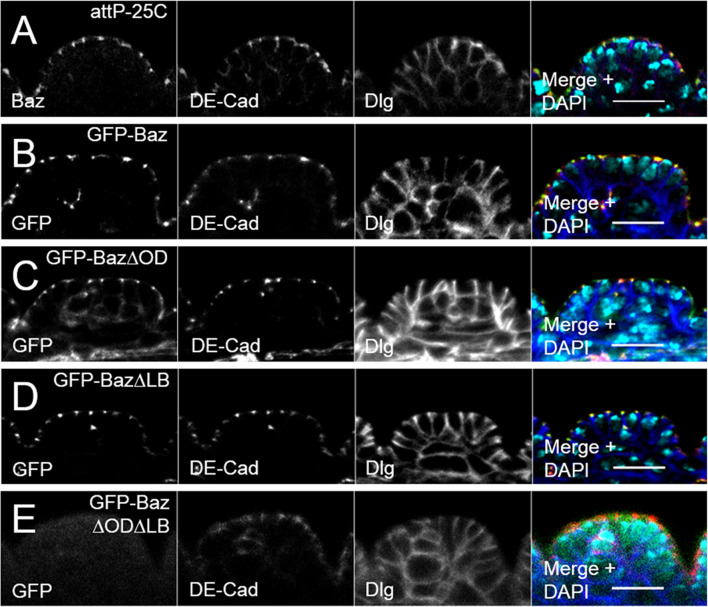
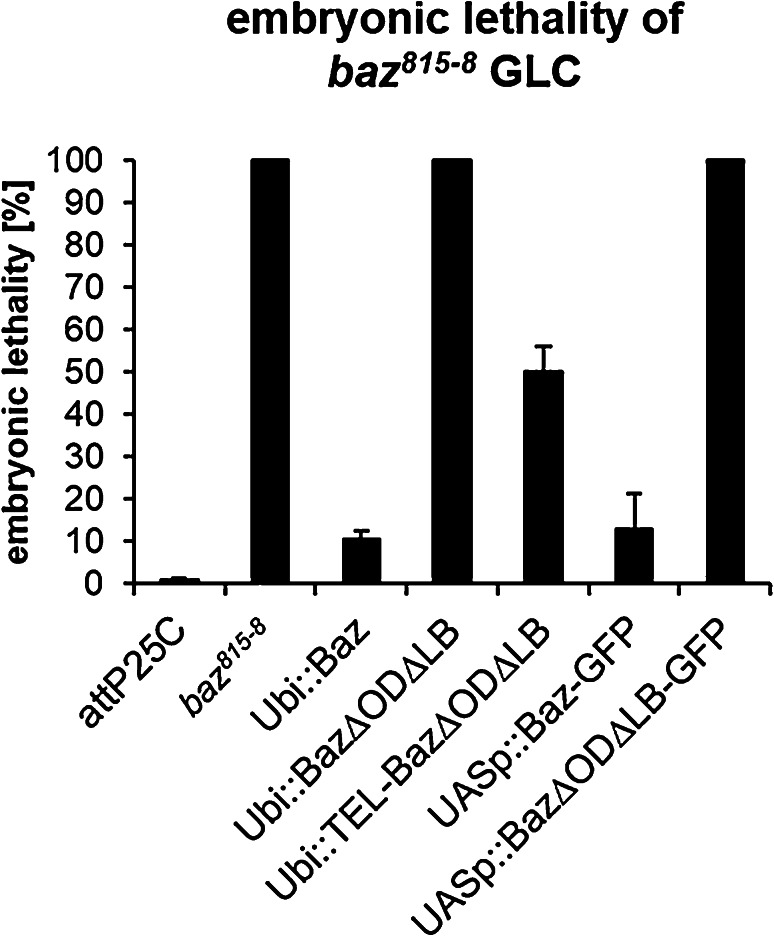
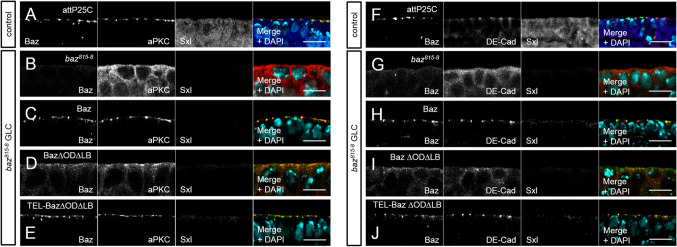
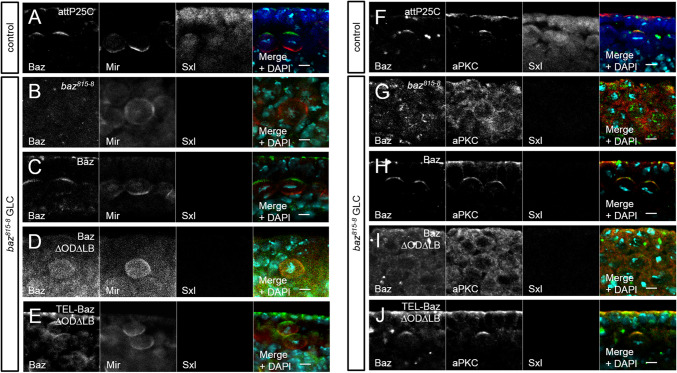
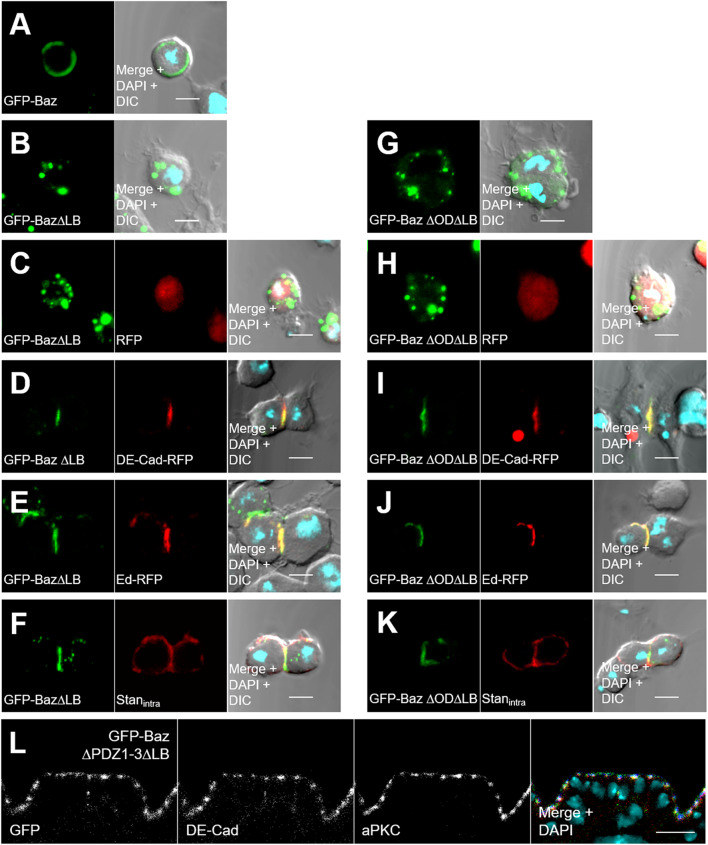
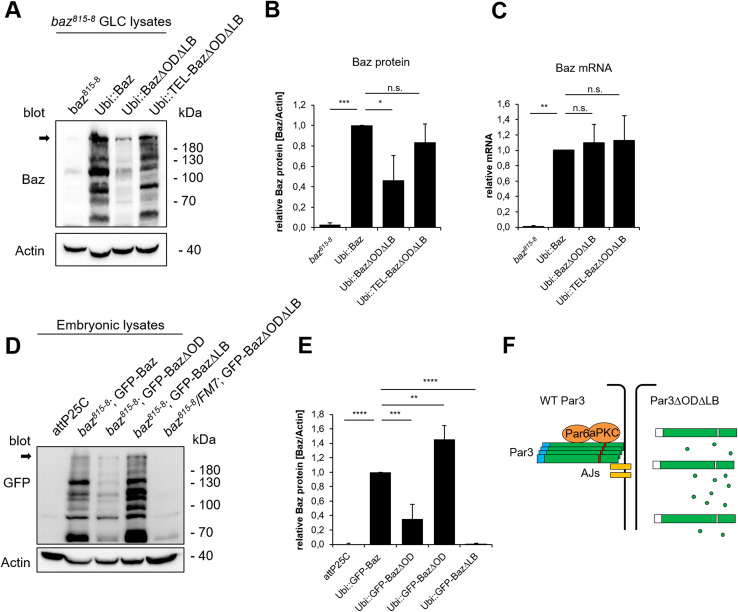
Apical-basal polarity is an important characteristic of epithelia and Drosophila neural stem cells. The conserved Par complex, which consists of the atypical protein kinase C and the scaffold proteins Baz and Par6, is a key player in the establishment of apical-basal cell polarity. Membrane recruitment of Baz has been reported to be accomplished by several mechanisms, which might function in redundancy, to ensure the correct localization of the complex. However, none of the described interactions was sufficient to displace the protein from the apical junctions. Here, we dissected the role of the oligomerization domain and the lipid-binding motif of Baz in vivo in the Drosophila embryo. We found that these domains function in redundancy to ensure the apical junctional localization of Baz: inactivation of only one domain is not sufficient to disrupt the function of Baz during apical-basal polarization of epithelial cells and neural stem cells. In contrast, mutation of both domains results in a strongly impaired protein stability and a phenotype characterized by embryonic lethality and an impaired apical-basal polarity in the embryonic epithelium and neural stem cells, resembling a baz-loss of function allele. Strikingly, the binding of Baz to the transmembrane proteins E-Cadherin, Echinoid, and Starry Night was not affected in this mutant protein. Our findings reveal a redundant function of the oligomerization and the lipid-binding domain, which is required for protein stability, correct subcellular localization, and apical-basal cell polarization.
Keywords: Adherens junctions; Cell polarity; Drosophila; Lipid binding; Par3.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Par-3 family proteins in cell polarity & adhesion.FEBS J. 2022 Feb;289(3):596-613. doi: 10.1111/febs.15754. Epub 2021 Mar 3. FEBS J. 2022. PMID: 33565714 Free PMC article. Review.
-
Rap1 acts via multiple mechanisms to position Canoe and adherens junctions and mediate apical-basal polarity establishment.Development. 2018 Jan 26;145(2):dev157941. doi: 10.1242/dev.157941. Development. 2018. PMID: 29361565 Free PMC article.
-
aPKC Inhibition by Par3 CR3 Flanking Regions Controls Substrate Access and Underpins Apical-Junctional Polarization.Dev Cell. 2016 Aug 22;38(4):384-98. doi: 10.1016/j.devcel.2016.07.018. Dev Cell. 2016. PMID: 27554858 Free PMC article.
-
Assembly of Bazooka polarity landmarks through a multifaceted membrane-association mechanism.J Cell Sci. 2012 Mar 1;125(Pt 5):1177-90. doi: 10.1242/jcs.091884. Epub 2012 Feb 2. J Cell Sci. 2012. PMID: 22303000
-
The apical polarity protein network in Drosophila epithelial cells: regulation of polarity, junctions, morphogenesis, cell growth, and survival.Annu Rev Cell Dev Biol. 2012;28:655-85. doi: 10.1146/annurev-cellbio-092910-154033. Epub 2012 Aug 6. Annu Rev Cell Dev Biol. 2012. PMID: 22881460 Review.
Cited by
-
A CDK1 phosphorylation site on Drosophila PAR-3 regulates neuroblast polarisation and sensory organ formation.Elife. 2024 Jun 13;13:e97902. doi: 10.7554/eLife.97902. Elife. 2024. PMID: 38869055 Free PMC article.
-
Phospholipids of the Plasma Membrane - Regulators or Consequence of Cell Polarity?Front Cell Dev Biol. 2020 Apr 28;8:277. doi: 10.3389/fcell.2020.00277. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 32411703 Free PMC article. Review.
-
PI(4,5)P2 controls slit diaphragm formation and endocytosis in Drosophila nephrocytes.Cell Mol Life Sci. 2022 Apr 18;79(5):248. doi: 10.1007/s00018-022-04273-7. Cell Mol Life Sci. 2022. PMID: 35437696 Free PMC article.
-
Par-3 family proteins in cell polarity & adhesion.FEBS J. 2022 Feb;289(3):596-613. doi: 10.1111/febs.15754. Epub 2021 Mar 3. FEBS J. 2022. PMID: 33565714 Free PMC article. Review.
-
Apical-basal polarity regulators are essential for slit diaphragm assembly and endocytosis in Drosophila nephrocytes.Cell Mol Life Sci. 2021 Apr;78(7):3657-3672. doi: 10.1007/s00018-021-03769-y. Epub 2021 Mar 2. Cell Mol Life Sci. 2021. PMID: 33651172 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
