

Innate Antiviral Immunity in the Skin
- PMID: 29526487
- PMCID: PMC5993211
- DOI: 10.1016/j.it.2018.02.003
Innate Antiviral Immunity in the Skin
Abstract
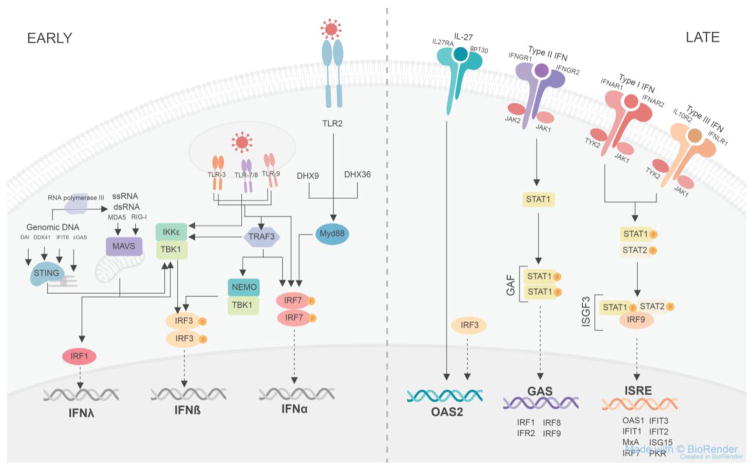
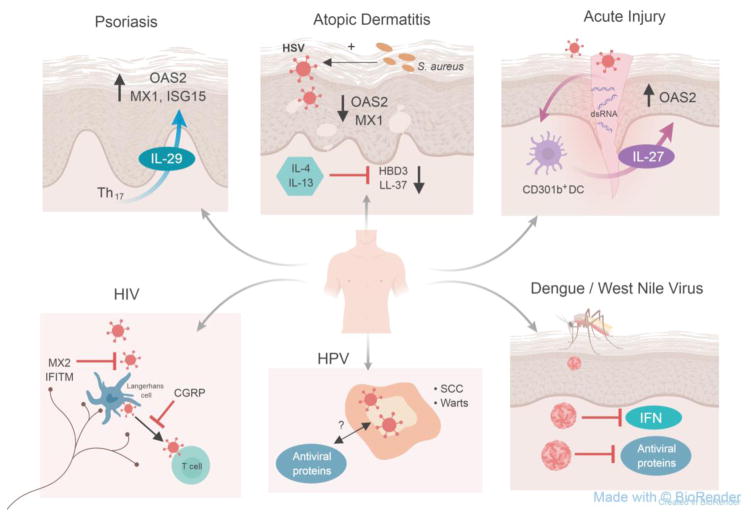
Barrier sites such as the skin play a critical role in immune defense. They must maintain homeostasis with commensals and rapidly detect and limit pathogen invasion. This is accomplished in part through the production of endogenous antimicrobial peptides and proteins, which can be either constitutive or inducible. Here, we focus particularly on the control of innate antiviral proteins and present the basic aspects of their regulation in the skin by interferons (IFNs), IFN-independent immunity, and environmental factors. We also discuss the activity and (dys-)function of antiviral proteins in the context of skin-tropic viruses and highlight the relevance of the innate antiviral pathway as a potential therapeutic avenue for vulnerable patient populations and skin diseases with high risk for virus infections.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
