

Combined Loss of JMJD1A and JMJD1B Reveals Critical Roles for H3K9 Demethylation in the Maintenance of Embryonic Stem Cells and Early Embryogenesis
- PMID: 29526734
- PMCID: PMC5998703
- DOI: 10.1016/j.stemcr.2018.02.002
Combined Loss of JMJD1A and JMJD1B Reveals Critical Roles for H3K9 Demethylation in the Maintenance of Embryonic Stem Cells and Early Embryogenesis
Abstract
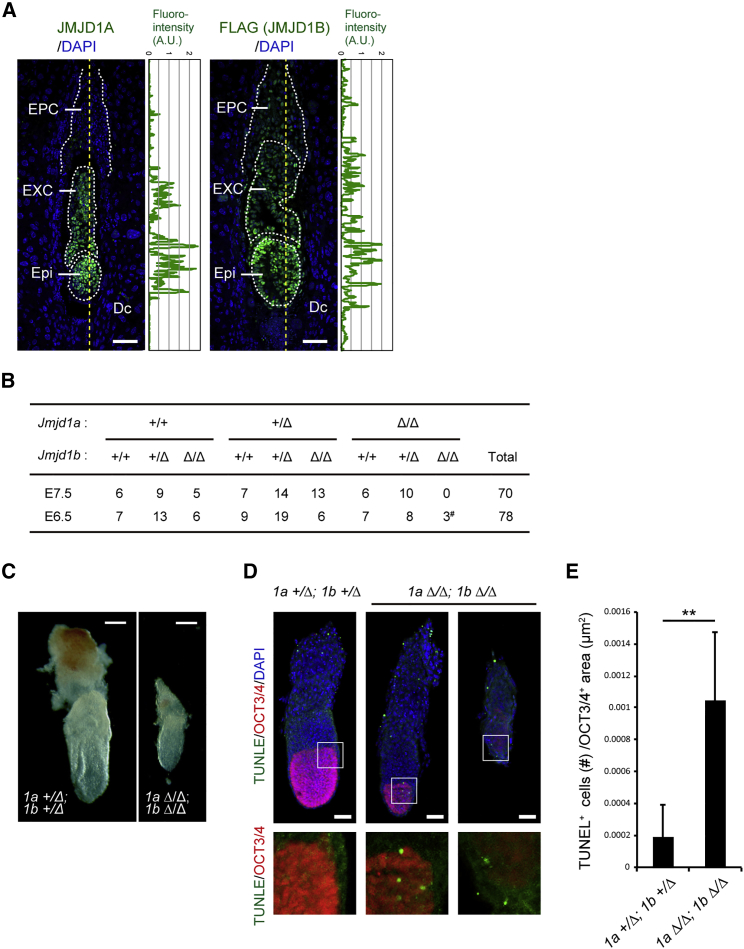
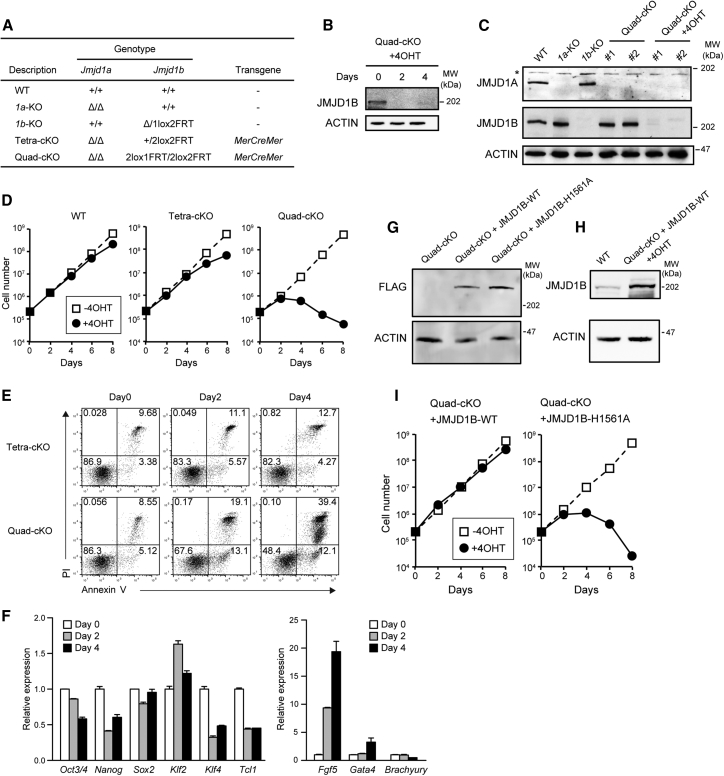
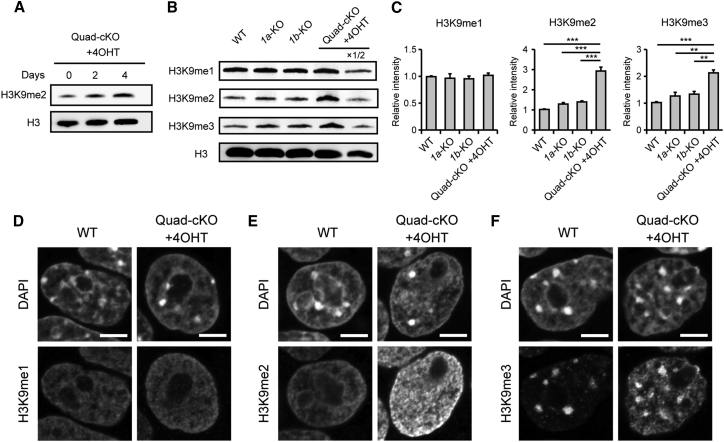
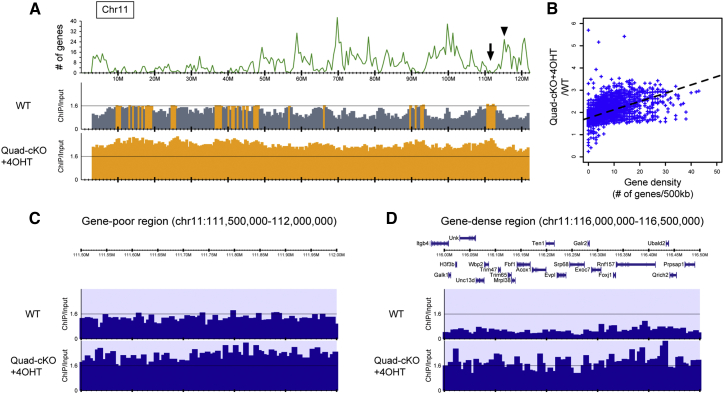
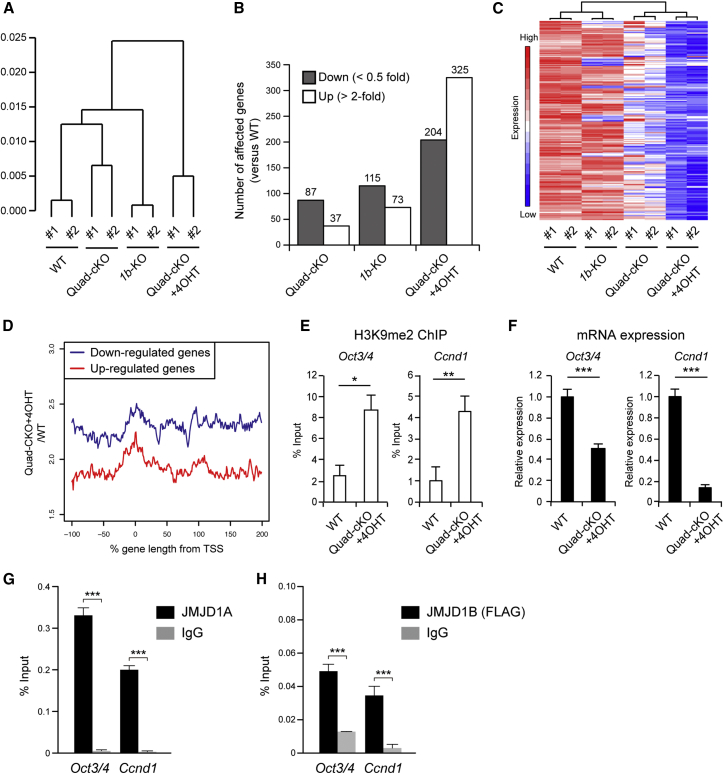
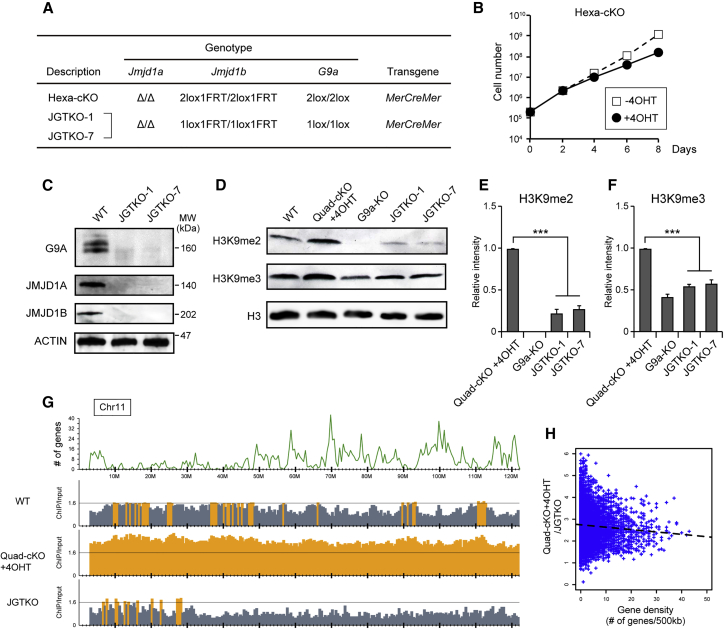
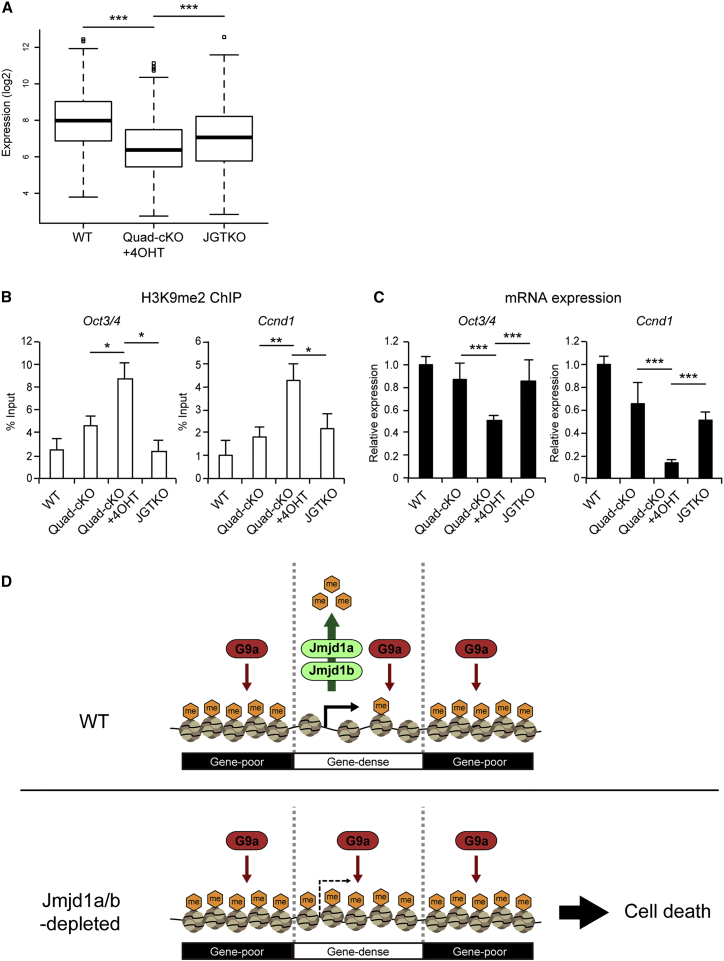
Histone H3 lysine 9 (H3K9) methylation is unevenly distributed in mammalian chromosomes. However, the molecular mechanism controlling the uneven distribution and its biological significance remain to be elucidated. Here, we show that JMJD1A and JMJD1B preferentially target H3K9 demethylation of gene-dense regions of chromosomes, thereby establishing an H3K9 hypomethylation state in euchromatin. JMJD1A/JMJD1B-deficient embryos died soon after implantation accompanying epiblast cell death. Furthermore, combined loss of JMJD1A and JMJD1B caused perturbed expression of metabolic genes and rapid cell death in embryonic stem cells (ESCs). These results indicate that JMJD1A/JMJD1B-meditated H3K9 demethylation has critical roles for early embryogenesis and ESC maintenance. Finally, genetic rescue experiments clarified that H3K9 overmethylation by G9A was the cause of the cell death and perturbed gene expression of JMJD1A/JMJD1B-depleted ESCs. We summarized that JMJD1A and JMJD1B, in combination, ensure early embryogenesis and ESC viability by establishing the correct H3K9 methylated epigenome.
Keywords: embryonic stem cell; histone demethylation; histone methylation; transcription.
Copyright © 2018 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Copeland N.G., Jenkins N.A., Court D.L. Recombineering: a powerful new tool for mouse functional genomics. Nat. Rev. Genet. 2001;2:769–779. - PubMed
-
- Feldman N., Gerson A., Fang J., Li E., Zhang Y., Shinkai Y., Cedar H., Bergman Y. G9a-mediated irreversible epigenetic inactivation of Oct-3/4 during early embryogenesis. Nat. Cell Biol. 2006;8:188–194. - PubMed
-
- Ferry L., Fournier A., Tsusaka T., Adelmant G., Shimazu T., Matano S., Kirsh O., Amouroux R., Dohmae N., Suzuki T. Methylation of DNA ligase 1 by G9a/GLP recruits UHRF1 to replicating DNA and regulates DNA methylation. Mol. Cell. 2017;67:550–565.e5. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
