

Müller glia-derived PRSS56 is required to sustain ocular axial growth and prevent refractive error
- PMID: 29529029
- PMCID: PMC5864079
- DOI: 10.1371/journal.pgen.1007244
Müller glia-derived PRSS56 is required to sustain ocular axial growth and prevent refractive error
Abstract
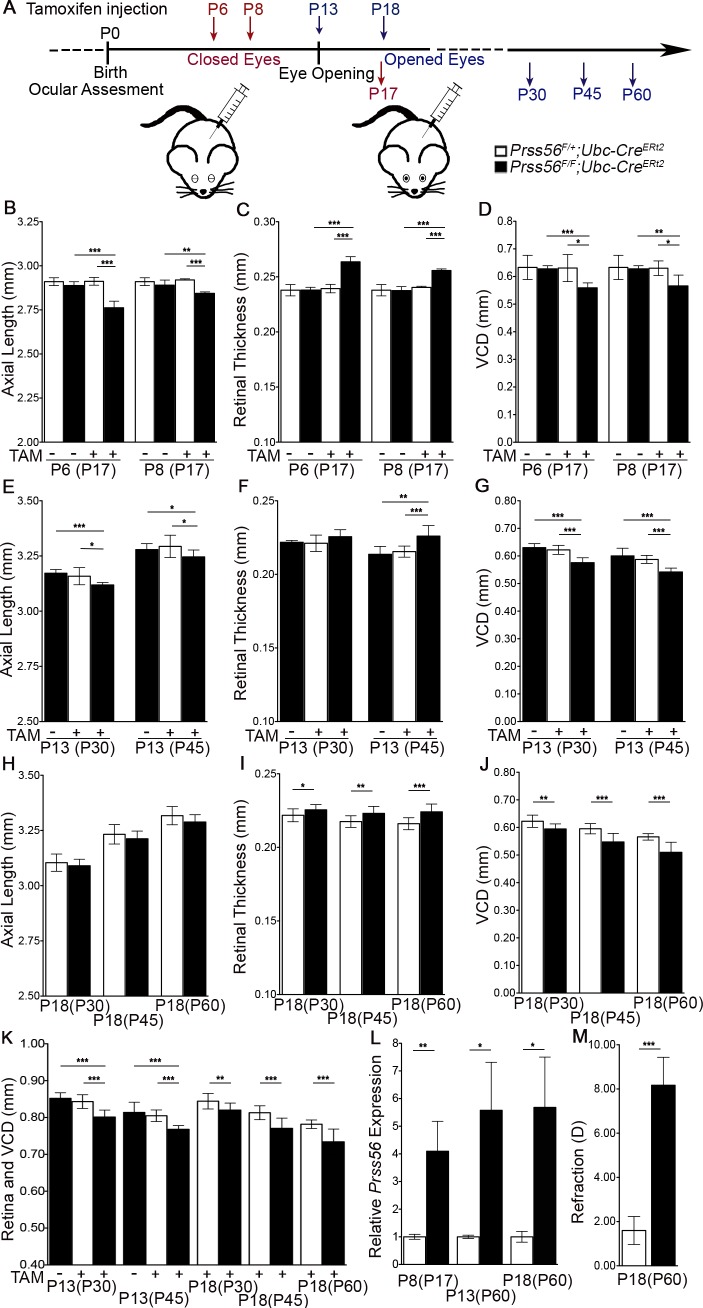
A mismatch between optical power and ocular axial length results in refractive errors. Uncorrected refractive errors constitute the most common cause of vision loss and second leading cause of blindness worldwide. Although the retina is known to play a critical role in regulating ocular growth and refractive development, the precise factors and mechanisms involved are poorly defined. We have previously identified a role for the secreted serine protease PRSS56 in ocular size determination and PRSS56 variants have been implicated in the etiology of both hyperopia and myopia, highlighting its importance in refractive development. Here, we use a combination of genetic mouse models to demonstrate that Prss56 mutations leading to reduced ocular size and hyperopia act via a loss of function mechanism. Using a conditional gene targeting strategy, we show that PRSS56 derived from Müller glia contributes to ocular growth, implicating a new retinal cell type in ocular size determination. Importantly, we demonstrate that persistent activity of PRSS56 is required during distinct developmental stages spanning the pre- and post-eye opening periods to ensure optimal ocular growth. Thus, our mouse data provide evidence for the existence of a molecule contributing to both the prenatal and postnatal stages of human ocular growth. Finally, we demonstrate that genetic inactivation of Prss56 rescues axial elongation in a mouse model of myopia caused by a null mutation in Egr1. Overall, our findings identify PRSS56 as a potential therapeutic target for modulating ocular growth aimed at preventing or slowing down myopia, which is reaching epidemic proportions.
Conflict of interest statement
The authors have declared that no competing interests exist
Figures







References
-
- Stone RA, Pardue MT, Iuvone PM, Khurana TS. Pharmacology of myopia and potential role for intrinsic retinal circadian rhythms. Experimental eye research. 2013;114:35–47. Epub 2013/01/15. doi: 10.1016/j.exer.2013.01.001 ; PubMed Central PMCID: PMC3636148. - DOI - PMC - PubMed
-
- Meng W, Butterworth J, Malecaze F, Calvas P. Axial length of myopia: a review of current research. Ophthalmologica Journal international d'ophtalmologie International journal of ophthalmology Zeitschrift fur Augenheilkunde. 2011;225(3):127–34. Epub 2010/10/16. doi: 10.1159/000317072 . - DOI - PubMed
-
- Bourne RR, Stevens GA, White RA, Smith JL, Flaxman SR, Price H, et al. Causes of vision loss worldwide, 1990–2010: a systematic analysis. The Lancet Global health. 2013;1(6):e339–49. Epub 2014/08/12. doi: 10.1016/S2214-109X(13)70113-X . - DOI - PubMed
-
- Dolgin E. The myopia boom. Nature. 2015;519(7543):276–8. Epub 2015/03/20. doi: 10.1038/519276a . - DOI - PubMed
-
- Tarutta E, Chua WH, Young T, Goldschmidt E, Saw SM, Rose KA, et al. Myopia: Why Study the Mechanisms of Myopia? Novel Approaches to Risk Factors Signalling Eye Growth- How Could Basic Biology Be Translated into Clinical Insights? Where Are Genetic and Proteomic Approaches Leading? How Does Visual Function Contribute to and Interact with Ametropia? Does Eye Shape Matter? Why Ametropia at All? Optometry and vision science: official publication of the American Academy of Optometry. 2011. Epub 2011/02/08. doi: 10.1097/OPX.0b013e31820e6a6a . - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
