

Host cell cytosolic immune response during Plasmodium liver stage development
- PMID: 29529207
- PMCID: PMC5995216
- DOI: 10.1093/femsre/fuy007
Host cell cytosolic immune response during Plasmodium liver stage development
Abstract
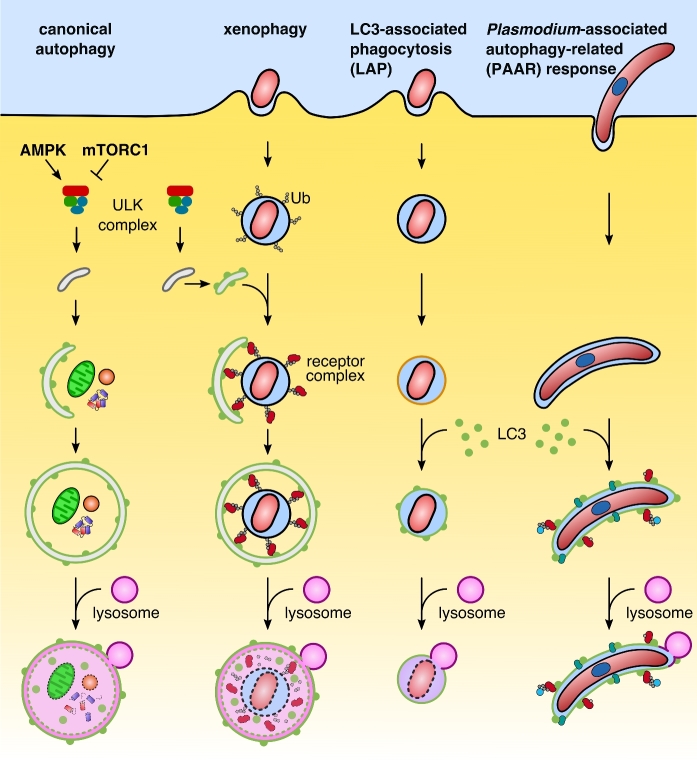
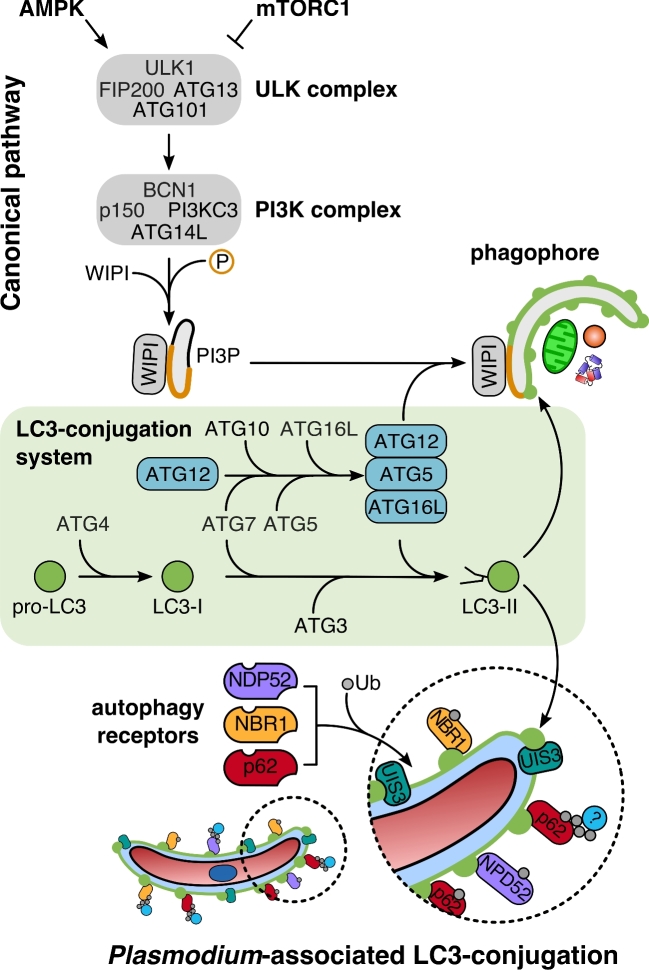
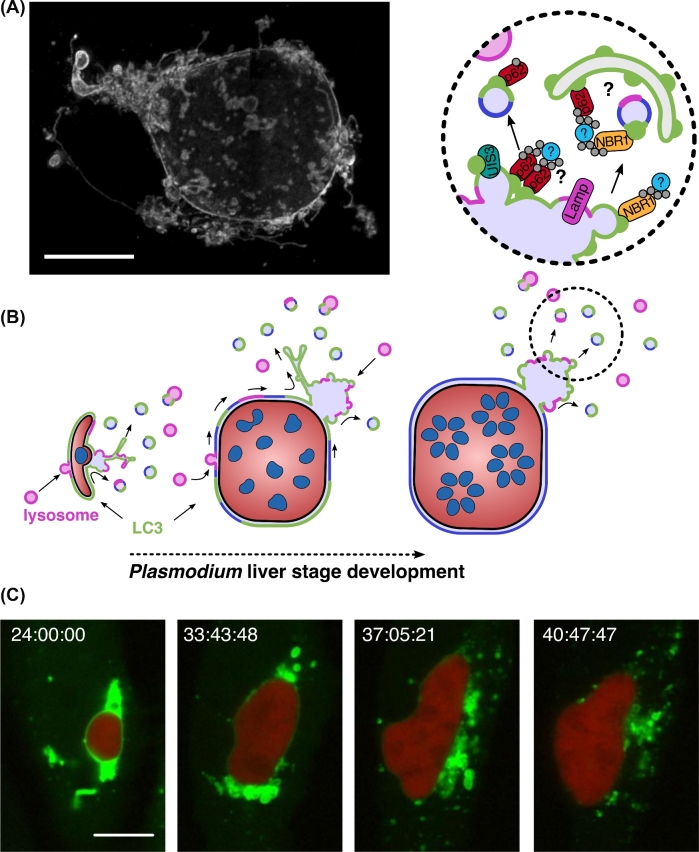
Recent years have witnessed a great gain in knowledge regarding parasite-host cell interactions during Plasmodium liver stage development. It is now an accepted fact that a large percentage of sporozoites invading hepatocytes fail to form infectious merozoites. There appears to be a delicate balance between parasite survival and elimination and we now start to understand why this is so. Plasmodium liver stage parasites replicate within the parasitophorous vacuole (PV), formed during invasion by invagination of the host cell plasma membrane. The main interface between the parasite and hepatocyte is the parasitophorous vacuole membrane (PVM) that surrounds the PV. Recently, it was shown that autophagy marker proteins decorate the PVM of Plasmodium liver stage parasites and eliminate a proportion of them by an autophagy-like mechanism. Successfully developing Plasmodium berghei parasites are initially also labeled but in the course of development, they are able to control this host defense mechanism by shedding PVM material into the tubovesicular network (TVN), an extension of the PVM that releases vesicles into the host cell cytoplasm. Better understanding of the molecular events at the PVM/TVN during parasite elimination could be the basis of new antimalarial measures.
Figures
Similar articles
-
Long-term live imaging reveals cytosolic immune responses of host hepatocytes against Plasmodium infection and parasite escape mechanisms.Autophagy. 2015;11(9):1561-79. doi: 10.1080/15548627.2015.1067361. Autophagy. 2015. PMID: 26208778 Free PMC article.
-
Deciphering host lysosome-mediated elimination of Plasmodium berghei liver stage parasites.Sci Rep. 2019 May 28;9(1):7967. doi: 10.1038/s41598-019-44449-z. Sci Rep. 2019. PMID: 31138850 Free PMC article.
-
Shedding of host autophagic proteins from the parasitophorous vacuolar membrane of Plasmodium berghei.Sci Rep. 2017 May 19;7(1):2191. doi: 10.1038/s41598-017-02156-7. Sci Rep. 2017. PMID: 28526861 Free PMC article.
-
The Plasmodium liver-stage parasitophorous vacuole: A front-line of communication between parasite and host.Int J Med Microbiol. 2018 Jan;308(1):107-117. doi: 10.1016/j.ijmm.2017.09.008. Epub 2017 Sep 15. Int J Med Microbiol. 2018. PMID: 28964681 Review.
-
Molecular make-up of the Plasmodium parasitophorous vacuolar membrane.Int J Med Microbiol. 2012 Oct;302(4-5):179-86. doi: 10.1016/j.ijmm.2012.07.011. Epub 2012 Aug 13. Int J Med Microbiol. 2012. PMID: 22898489 Review.
Cited by
-
Guidelines for the use and interpretation of assays for monitoring autophagy (4th edition)1.Autophagy. 2021 Jan;17(1):1-382. doi: 10.1080/15548627.2020.1797280. Epub 2021 Feb 8. Autophagy. 2021. PMID: 33634751 Free PMC article.
-
Hardly Vacuous: The Parasitophorous Vacuolar Membrane of Malaria Parasites.Trends Parasitol. 2020 Feb;36(2):138-146. doi: 10.1016/j.pt.2019.11.006. Epub 2019 Dec 19. Trends Parasitol. 2020. PMID: 31866184 Free PMC article. Review.
-
Plasmodium's fight for survival: escaping elimination while acquiring nutrients.Trends Parasitol. 2022 Jul;38(7):544-557. doi: 10.1016/j.pt.2022.04.004. Epub 2022 May 6. Trends Parasitol. 2022. PMID: 35534377 Free PMC article. Review.
-
Host-pathogen interactions in the Plasmodium-infected mouse liver at spatial and single-cell resolution.Nat Commun. 2024 Aug 19;15(1):7105. doi: 10.1038/s41467-024-51418-2. Nat Commun. 2024. PMID: 39160174 Free PMC article.
-
Mid-Liver Stage Arrest of Plasmodium falciparum Schizonts in Primary Porcine Hepatocytes.Front Cell Infect Microbiol. 2022 Feb 17;12:834850. doi: 10.3389/fcimb.2022.834850. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 35252038 Free PMC article.
References
-
- Amino R, Thiberge S, Martin B et al. . Quantitative imaging of Plasmodium transmission from mosquito to mammal. Nat Med 2006;12:220–4. - PubMed
-
- Annoura T, van Schaijk BCL, Ploemen IHJ et al. . Two Plasmodium 6-Cys family-related proteins have distinct and critical roles in liver-stage development. FASEB J 2014;28:2158–70. - PubMed
-
- Bano N, Romano JD, Jayabalasingham B et al. . Cellular interactions of Plasmodium liver stage with its host mammalian cell. Int J Parasitol 2007;37:1329–41. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
