

Protein composition of catalytically active U7-dependent processing complexes assembled on histone pre-mRNA containing biotin and a photo-cleavable linker
- PMID: 29529248
- PMCID: PMC5961079
- DOI: 10.1093/nar/gky133
Protein composition of catalytically active U7-dependent processing complexes assembled on histone pre-mRNA containing biotin and a photo-cleavable linker
Abstract
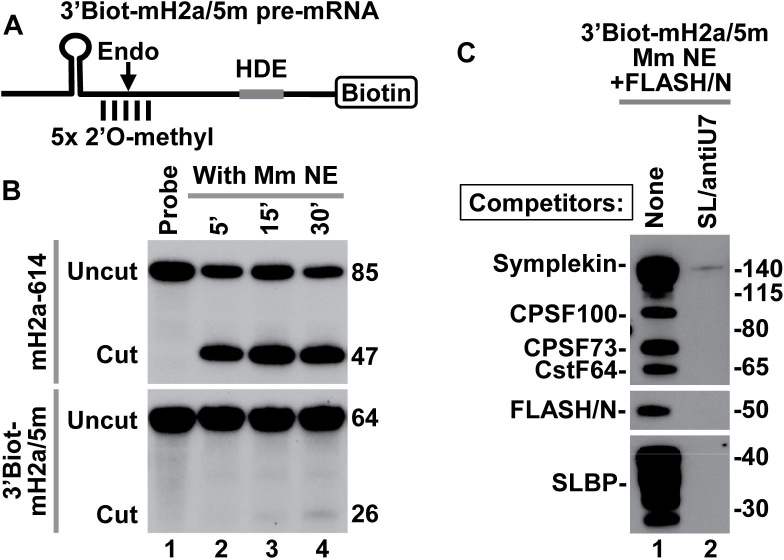
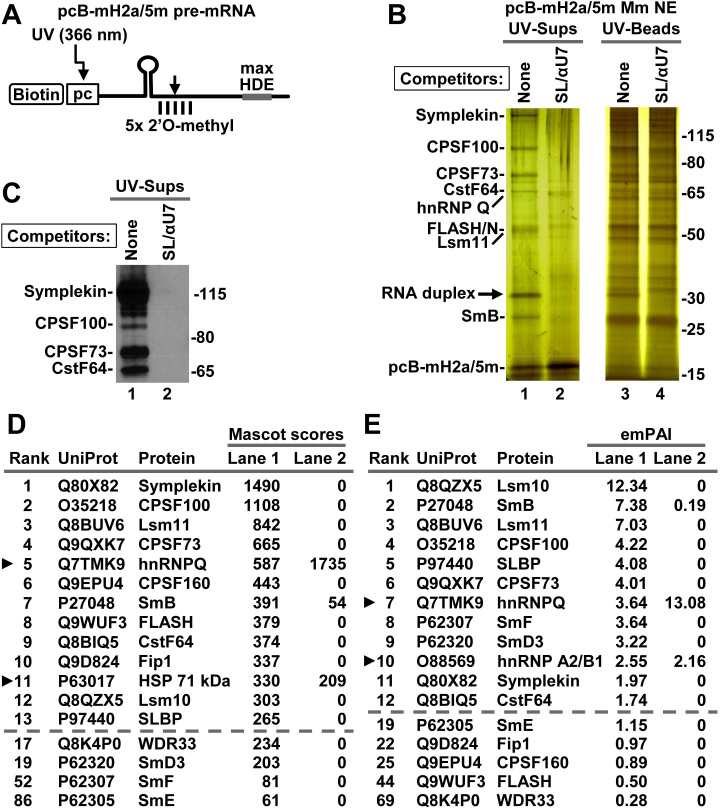
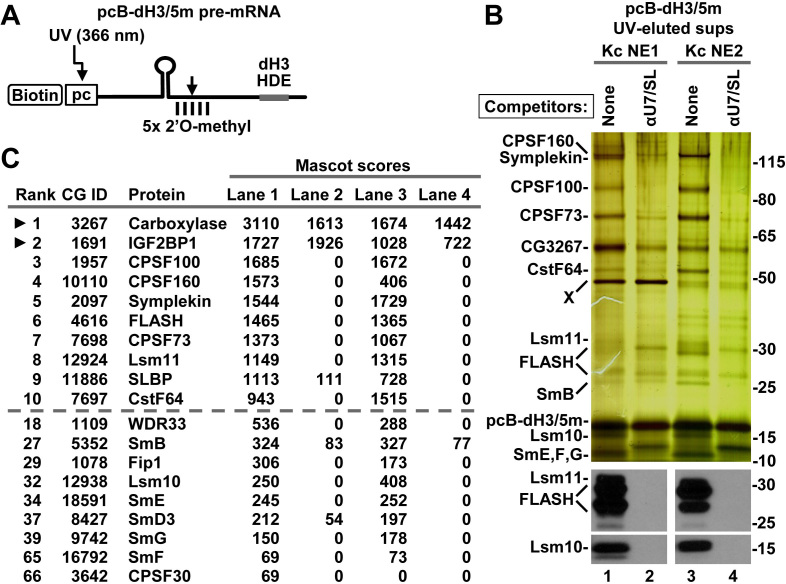
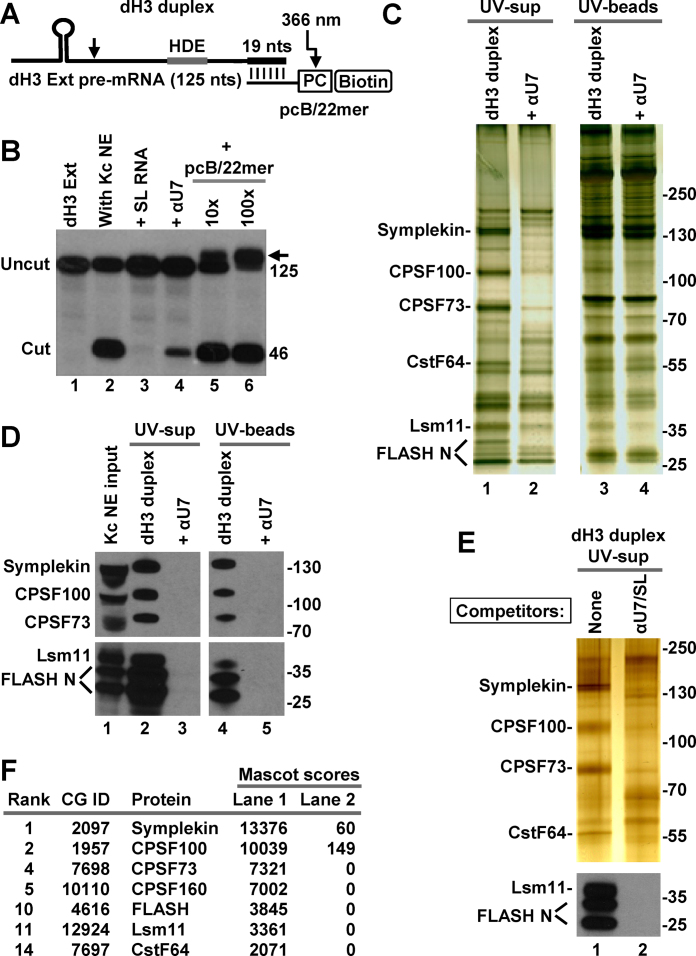
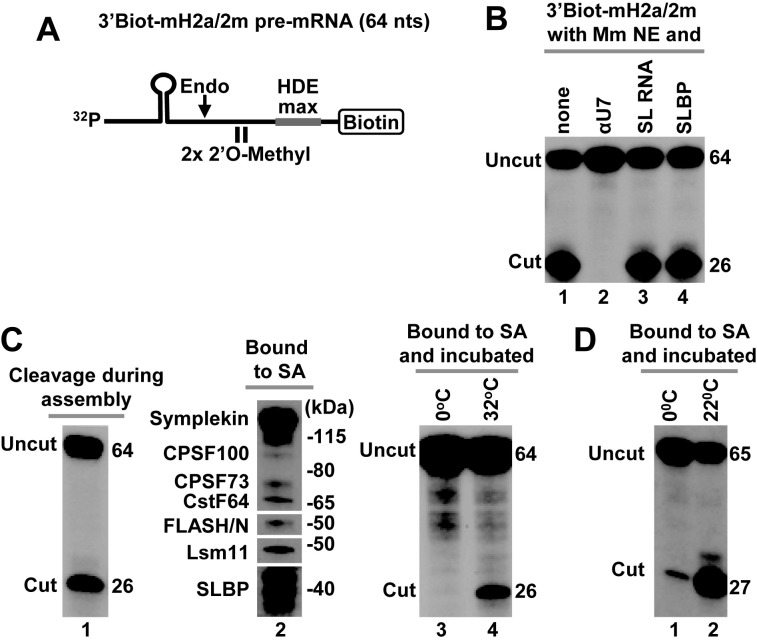
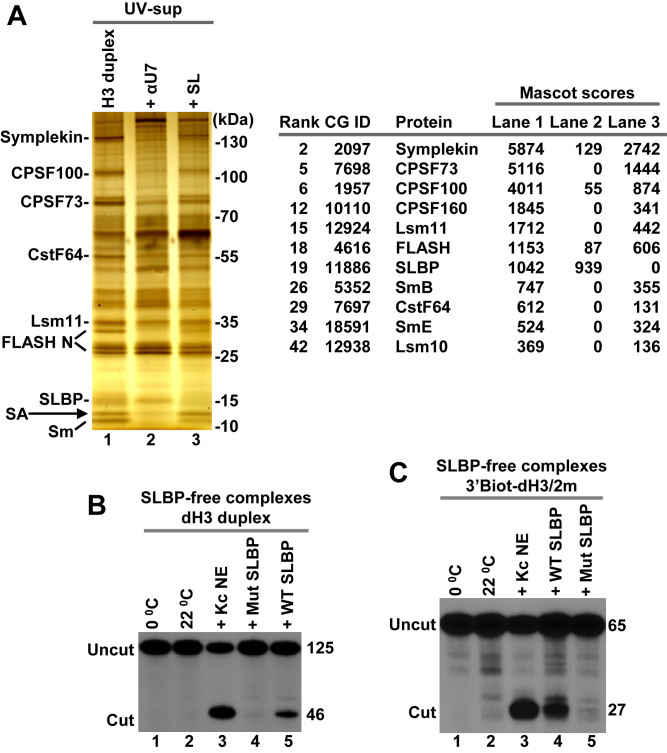
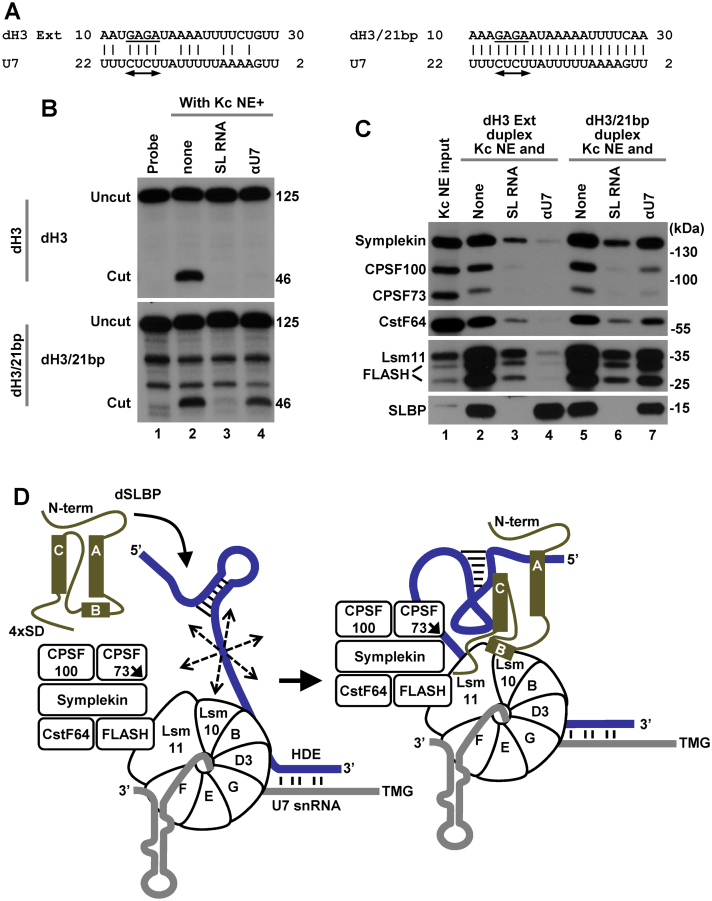
3' end cleavage of metazoan replication-dependent histone pre-mRNAs requires the multi-subunit holo-U7 snRNP and the stem-loop binding protein (SLBP). The exact composition of the U7 snRNP and details of SLBP function in processing remain unclear. To identify components of the U7 snRNP in an unbiased manner, we developed a novel approach for purifying processing complexes from Drosophila and mouse nuclear extracts. In this method, catalytically active processing complexes are assembled in vitro on a cleavage-resistant histone pre-mRNA containing biotin and a photo-sensitive linker, and eluted from streptavidin beads by UV irradiation for direct analysis by mass spectrometry. In the purified processing complexes, Drosophila and mouse U7 snRNP have a remarkably similar composition, always being associated with CPSF73, CPSF100, symplekin and CstF64. Many other proteins previously implicated in the U7-dependent processing are not present. Drosophila U7 snRNP bound to histone pre-mRNA in the absence of SLBP contains the same subset of polyadenylation factors but is catalytically inactive and addition of recombinant SLBP is sufficient to trigger cleavage. This result suggests that Drosophila SLBP promotes a structural rearrangement of the processing complex, resulting in juxtaposition of the CPSF73 endonuclease with the cleavage site in the pre-mRNA substrate.
Figures
Similar articles
-
3'-End processing of histone pre-mRNAs in Drosophila: U7 snRNP is associated with FLASH and polyadenylation factors.RNA. 2013 Dec;19(12):1726-44. doi: 10.1261/rna.040360.113. Epub 2013 Oct 21. RNA. 2013. PMID: 24145821 Free PMC article.
-
U7 snRNP is recruited to histone pre-mRNA in a FLASH-dependent manner by two separate regions of the stem-loop binding protein.RNA. 2017 Jun;23(6):938-951. doi: 10.1261/rna.060806.117. Epub 2017 Mar 13. RNA. 2017. PMID: 28289156 Free PMC article.
-
A complex containing the CPSF73 endonuclease and other polyadenylation factors associates with U7 snRNP and is recruited to histone pre-mRNA for 3'-end processing.Mol Cell Biol. 2013 Jan;33(1):28-37. doi: 10.1128/MCB.00653-12. Epub 2012 Oct 15. Mol Cell Biol. 2013. PMID: 23071092 Free PMC article.
-
U2 snRNP: not just for poly(A) mRNAs.Mol Cell. 2007 Nov 9;28(3):353-4. doi: 10.1016/j.molcel.2007.10.015. Mol Cell. 2007. PMID: 17996698 Review.
-
Formation of the 3' end of histone mRNA.Gene. 1999 Oct 18;239(1):1-14. doi: 10.1016/s0378-1119(99)00367-4. Gene. 1999. PMID: 10571029 Review.
Cited by
-
The CFII components PCF11 and Cbc change subnuclear localization as cells differentiate in the male germ line adult stem cell lineage.bioRxiv [Preprint]. 2025 Jul 22:2025.07.22.666077. doi: 10.1101/2025.07.22.666077. bioRxiv. 2025. Update in: MicroPubl Biol. 2025 Aug 15;2025. doi: 10.17912/micropub.biology.001718. PMID: 40777464 Free PMC article. Updated. Preprint.
-
Studies with recombinant U7 snRNP demonstrate that CPSF73 is both an endonuclease and a 5'-3' exonuclease.RNA. 2020 Oct;26(10):1345-1359. doi: 10.1261/rna.076273.120. Epub 2020 Jun 17. RNA. 2020. PMID: 32554553 Free PMC article.
-
Anticancer benzoxaboroles block pre-mRNA processing by directly inhibiting CPSF3.Cell Chem Biol. 2024 Jan 18;31(1):139-149.e14. doi: 10.1016/j.chembiol.2023.10.019. Epub 2023 Nov 14. Cell Chem Biol. 2024. PMID: 37967558 Free PMC article.
-
Superresolution light microscopy of the Drosophila histone locus body reveals a core-shell organization associated with expression of replication-dependent histone genes.Mol Biol Cell. 2021 Apr 19;32(9):942-955. doi: 10.1091/mbc.E20-10-0645. Epub 2021 Mar 31. Mol Biol Cell. 2021. PMID: 33788585 Free PMC article.
-
Structure of an active human histone pre-mRNA 3'-end processing machinery.Science. 2020 Feb 7;367(6478):700-703. doi: 10.1126/science.aaz7758. Science. 2020. PMID: 32029631 Free PMC article.
References
-
- Dominski Z., Marzluff W.F.. Three-hybrid screens for RNA-binding proteins: proteins binding the 3′ end of histone mRNA. Methods Mol. Biol. 2001; 177:291–318. - PubMed
-
- Marzluff W.F. Metazoan replication-dependent histone mRNAs: a distinct set of RNA polymerase II transcripts. Curr. Opin. Cell Biol. 2005; 17:274–280. - PubMed
-
- Dominski Z., Carpousis A.J., Clouet-d’Orval B.. Emergence of the beta-CASP ribonucleases: highly conserved and ubiquitous metallo-enzymes involved in messenger RNA maturation and degradation. Biochim. Biophys. Acta. 2013; 1829:532–551. - PubMed
-
- Galli G., Hofstetter H., Stunnenberg H.G., Birnstiel M.L.. Biochemical complementation with RNA in the Xenopus oocyte: a small RNA is required for the generation of 3′ histone mRNA termini. Cell. 1983; 34:823–828. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
