

Hepatitis E Virus Genome Structure and Replication Strategy
- PMID: 29530948
- PMCID: PMC6314074
- DOI: 10.1101/cshperspect.a031724
Hepatitis E Virus Genome Structure and Replication Strategy
Abstract
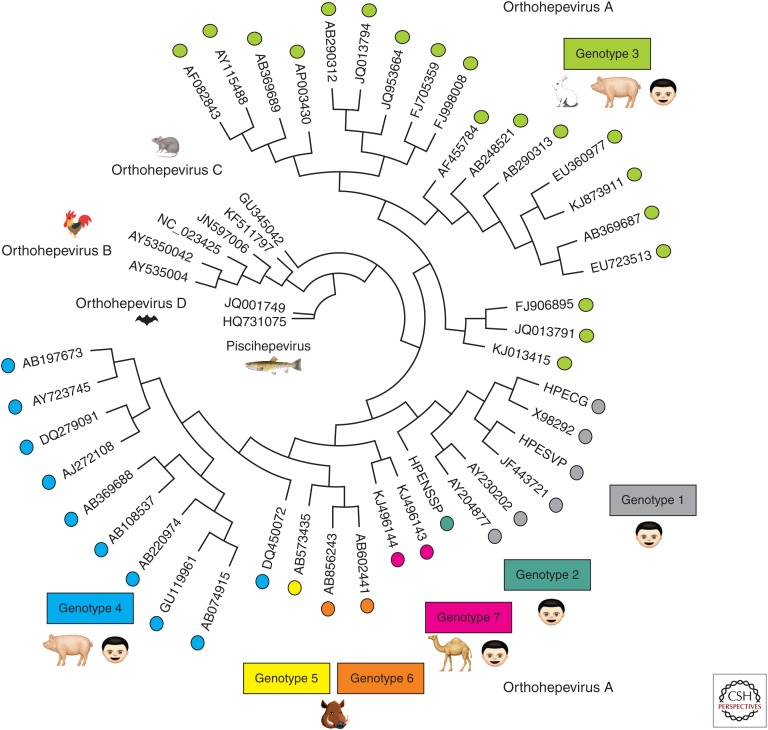
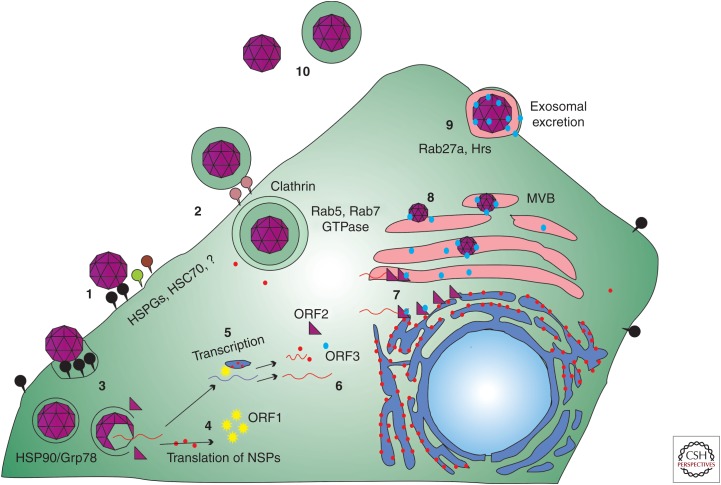
Hepatitis E virus (HEV) possesses many of the features of other positive-stranded RNA viruses but also adds HEV-specific nuances, making its virus-host interactions unique. Slow virus replication kinetics and fastidious growth conditions, coupled with the historical lack of an efficient cell culture system to propagate the virus, have left many gaps in our understanding of its structure and replication cycle. Recent advances in culturing selected strains of HEV and resolving the 3D structure of the viral capsid are filling in knowledge gaps, but HEV remains an extremely understudied pathogen. Many steps in the HEV life cycle and many aspects of HEV pathogenesis remain unknown, such as the host and viral factors that determine cross-species infection, the HEV-specific receptor(s) on host cells, what determines HEV chronicity and the ability to replicate in extrahepatic sites, and what regulates processing of the open reading frame 1 (ORF1) nonstructural polyprotein.
Copyright © 2019 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures

References
-
- Agrawal S, Gupta D, Panda SK. 2001. The 3′ end of hepatitis E virus (HEV) genome binds specifically to the viral RNA-dependent RNA polymerase (RdRp). Virology 282: 87–101. - PubMed
-
- Allen MD, Buckle AM, Cordell SC, Lowe J, Bycroft M. 2003. The crystal structure of AF1521 a protein from Archaeoglobus fulgidus with homology to the non-histone domain of macroH2A. J Mol Biol 330: 503–511. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources