

Immune evasion mediated by PD-L1 on glioblastoma-derived extracellular vesicles
- PMID: 29532035
- PMCID: PMC5842038
- DOI: 10.1126/sciadv.aar2766
Immune evasion mediated by PD-L1 on glioblastoma-derived extracellular vesicles
Abstract
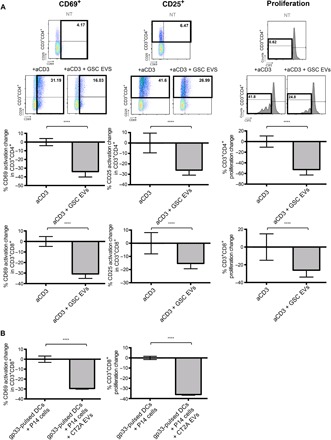
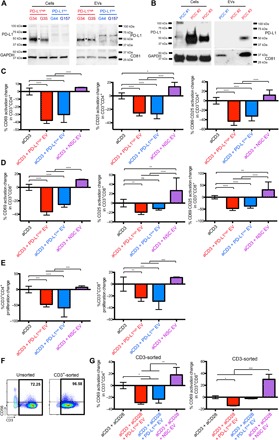
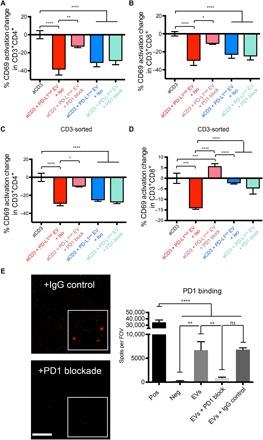
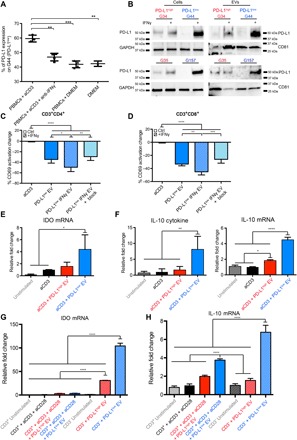
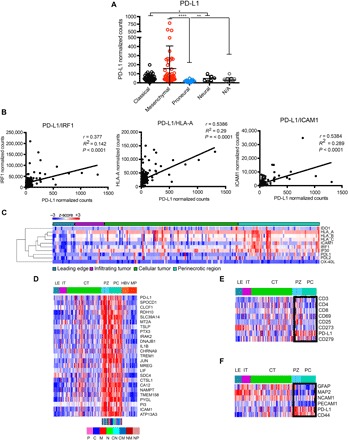
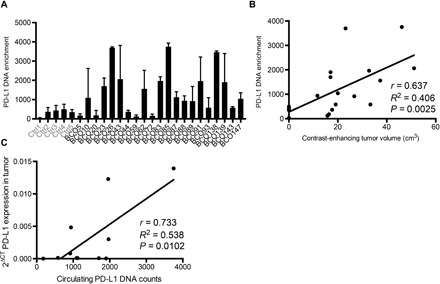
Binding of programmed death ligand-1 (PD-L1) to programmed cell death protein-1 (PD1) leads to cancer immune evasion via inhibition of T cell function. One of the defining characteristics of glioblastoma, a universally fatal brain cancer, is its profound local and systemic immunosuppression. Glioblastoma has also been shown to generate extracellular vesicles (EVs), which may play an important role in tumor progression. We thus hypothesized that glioblastoma EVs may be important mediators of immunosuppression and that PD-L1 could play a role. We show that glioblastoma EVs block T cell activation and proliferation in response to T cell receptor stimulation. PD-L1 was expressed on the surface of some, but not of all, glioblastoma-derived EVs, with the potential to directly bind to PD1. An anti-PD1 receptor blocking antibody significantly reversed the EV-mediated blockade of T cell activation but only when PD-L1 was present on EVs. When glioblastoma PD-L1 was up-regulated by IFN-γ, EVs also showed some PD-L1-dependent inhibition of T cell activation. PD-L1 expression correlated with the mesenchymal transcriptome profile and was anatomically localized in the perinecrotic and pseudopalisading niche of human glioblastoma specimens. PD-L1 DNA was present in circulating EVs from glioblastoma patients where it correlated with tumor volumes of up to 60 cm3. These results suggest that PD-L1 on EVs may be another mechanism for glioblastoma to suppress antitumor immunity and support the potential of EVs as biomarkers in tumor patients.
Figures
References
-
- Stupp R., Mason W. P., van den Bent M. J., Weller M., Fisher B., Taphoorn M. J. B., Belanger K., Brandes A. A., Marosi C., Bogdahn U., Curschmann J., Janzer R. C., Ludwin S. K., Gorlia T., Allgeier A., Lacombe D., Cairncross J. G., Eisenhauer E., Mirimanoff R. O. European Organisation for Research and Treatment of Cancer Brain Tumor and Radiotherapy Groups; National Cancer Institute of Canada Clinical Trials Group , Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 352, 987–996 (2005). - PubMed
-
- Mahoney K. M., Rennert P. D., Freeman G. J., Combination cancer immunotherapy and new immunomodulatory targets. Nat. Rev. Drug Discov. 14, 561–584 (2015). - PubMed
-
- Kmiecik J., Poli A., Brons N. H. C., Waha A., Eide G. E., Enger P. Ø., Zimmer J., Chekenya M., Elevated CD3+ and CD8+ tumor-infiltrating immune cells correlate with prolonged survival in glioblastoma patients despite integrated immunosuppressive mechanisms in the tumor microenvironment and at the systemic level. J. Neuroimmunol. 264, 71–83 (2013). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
