

Template-Independent Enzymatic Oligonucleotide Synthesis (TiEOS): Its History, Prospects, and Challenges
- PMID: 29533604
- PMCID: PMC7995564
- DOI: 10.1021/acs.biochem.7b00937
Template-Independent Enzymatic Oligonucleotide Synthesis (TiEOS): Its History, Prospects, and Challenges
Abstract
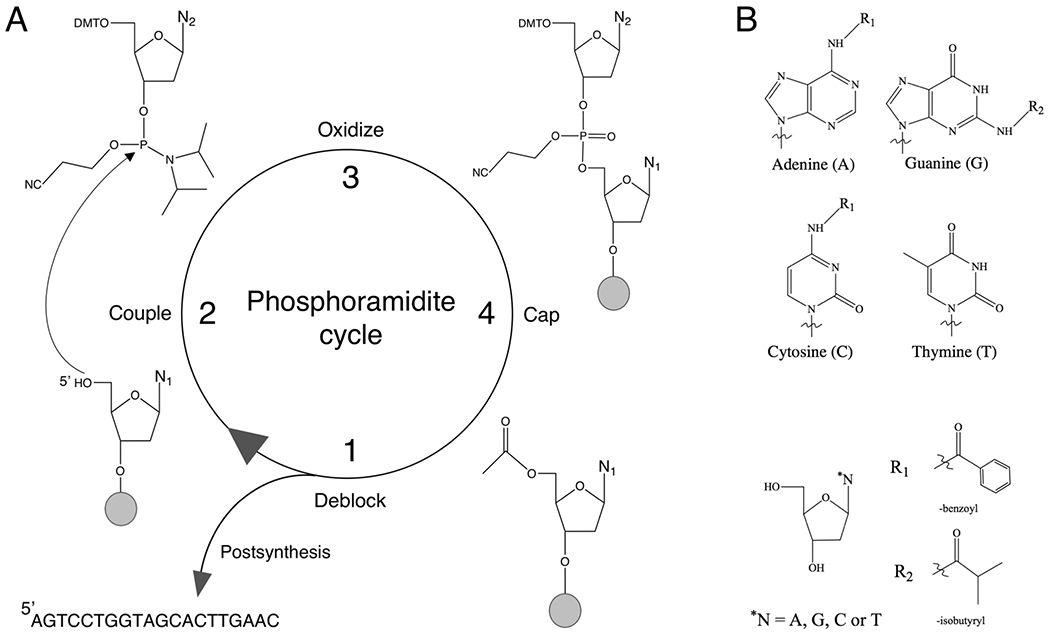
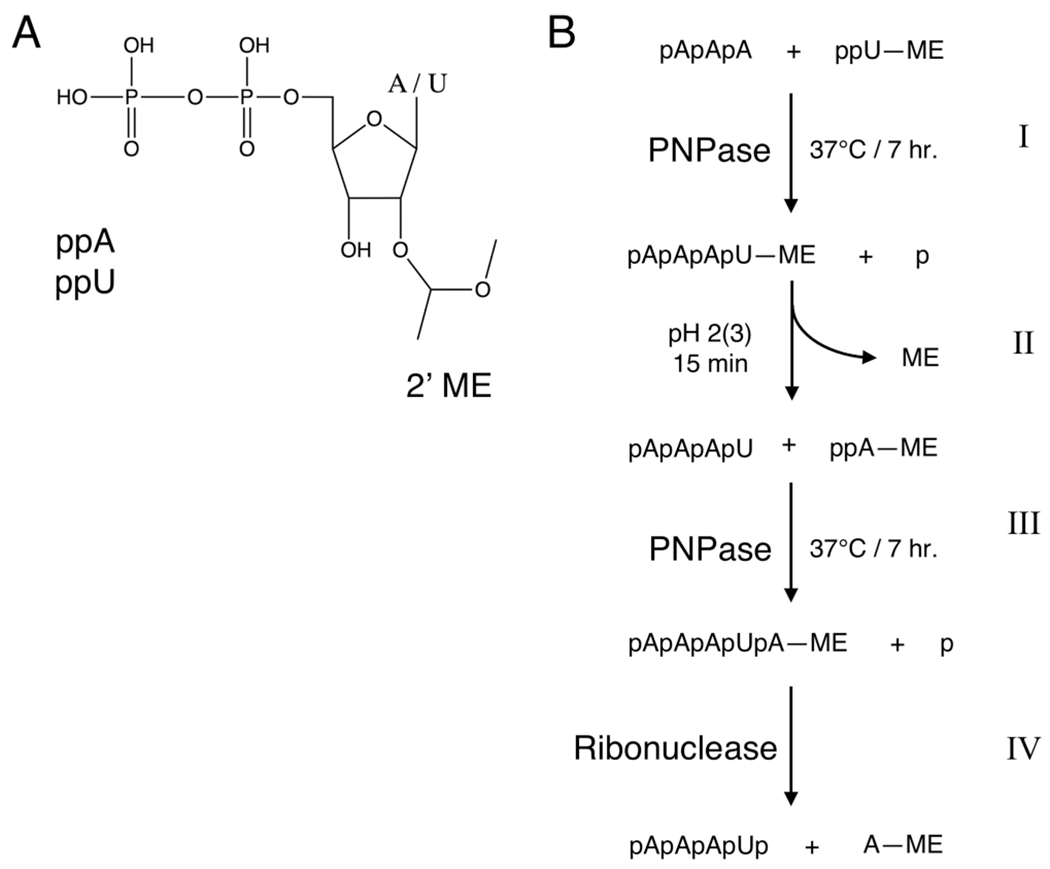
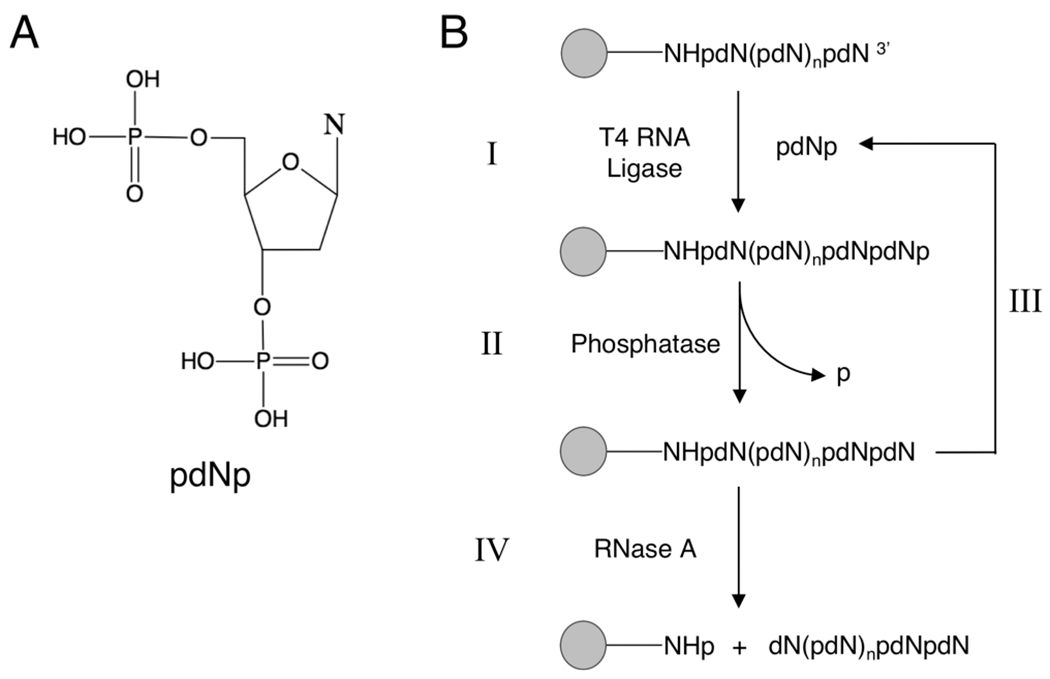
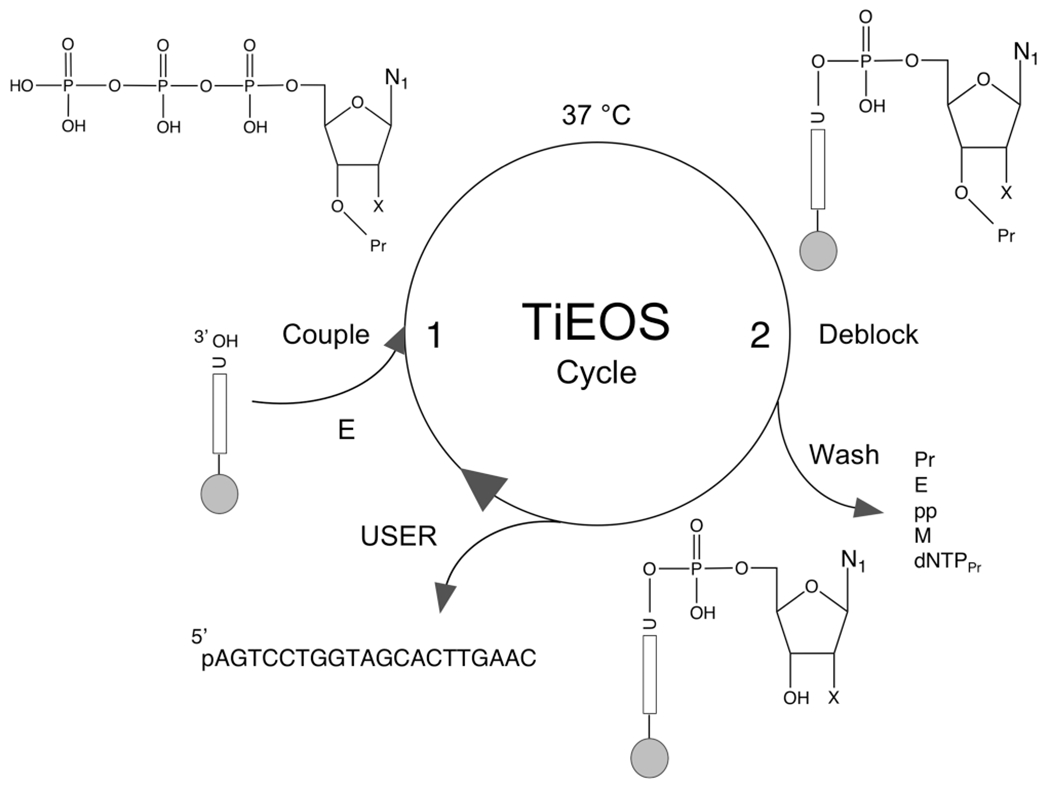
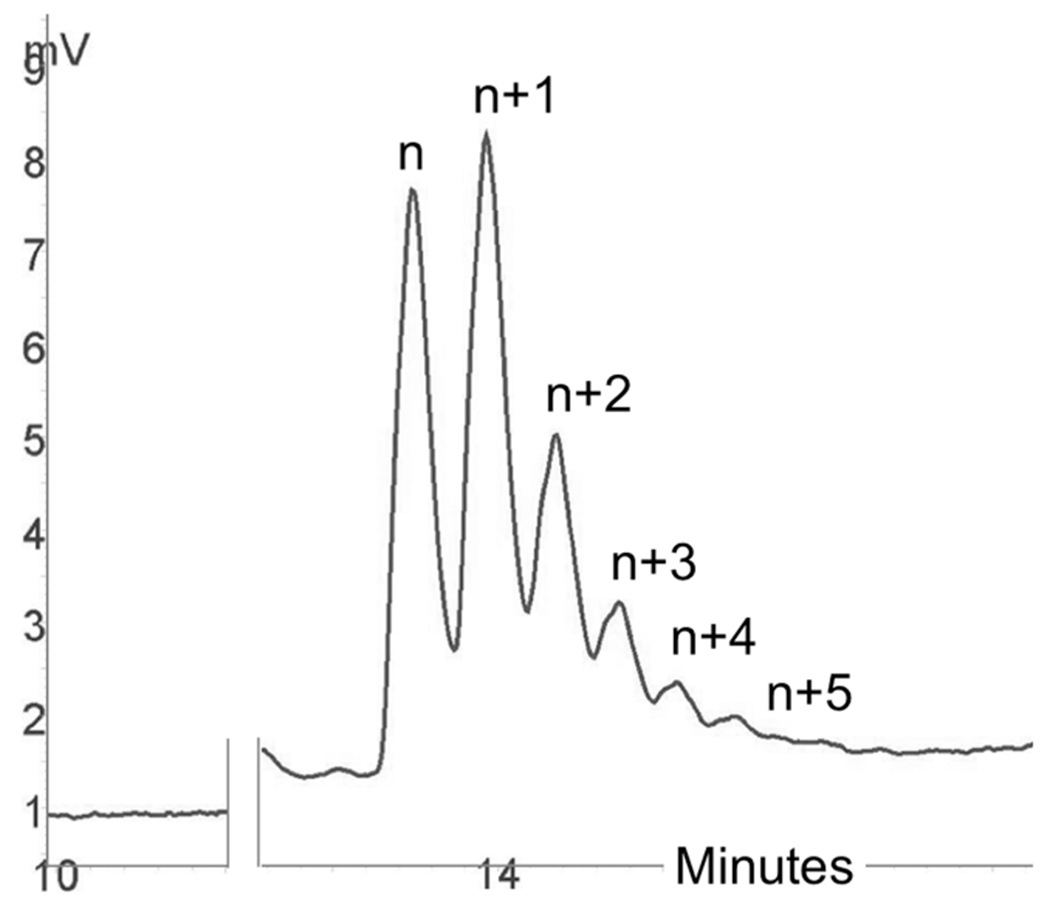
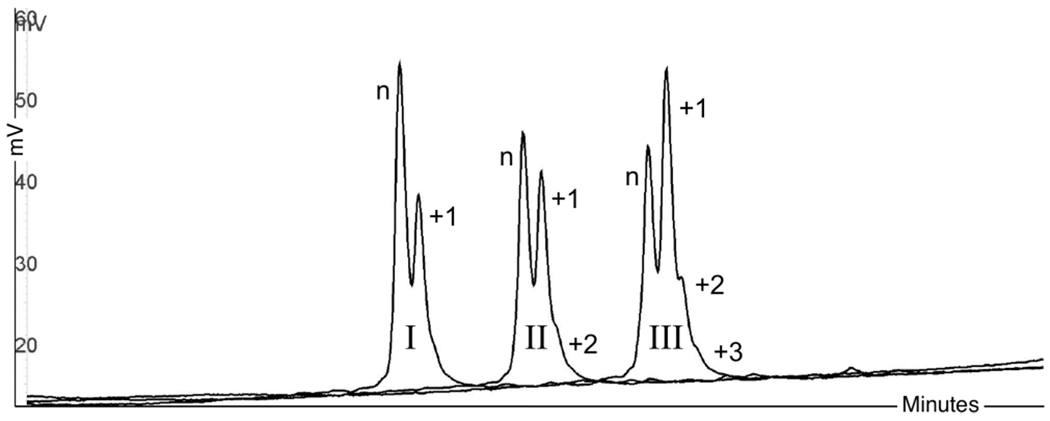
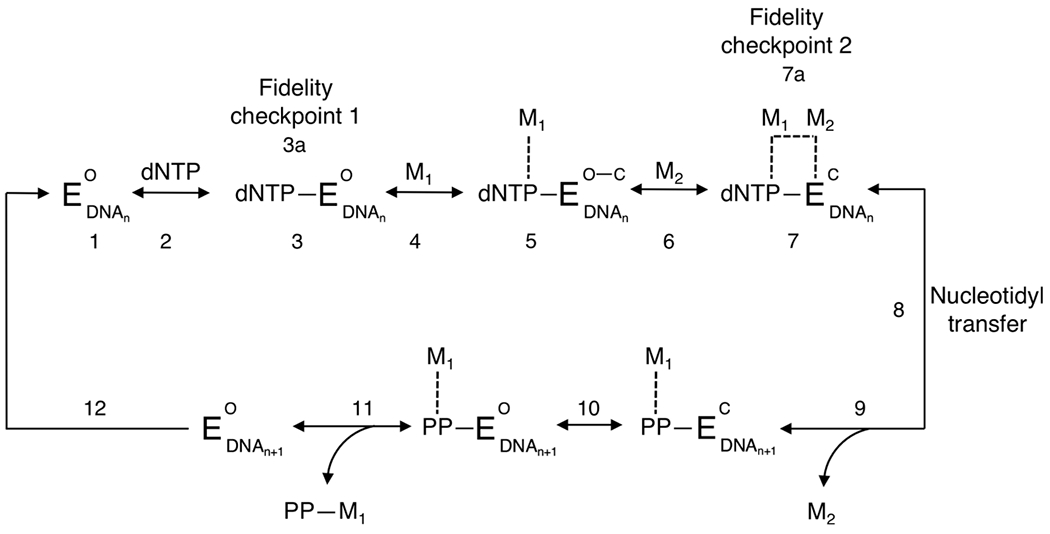
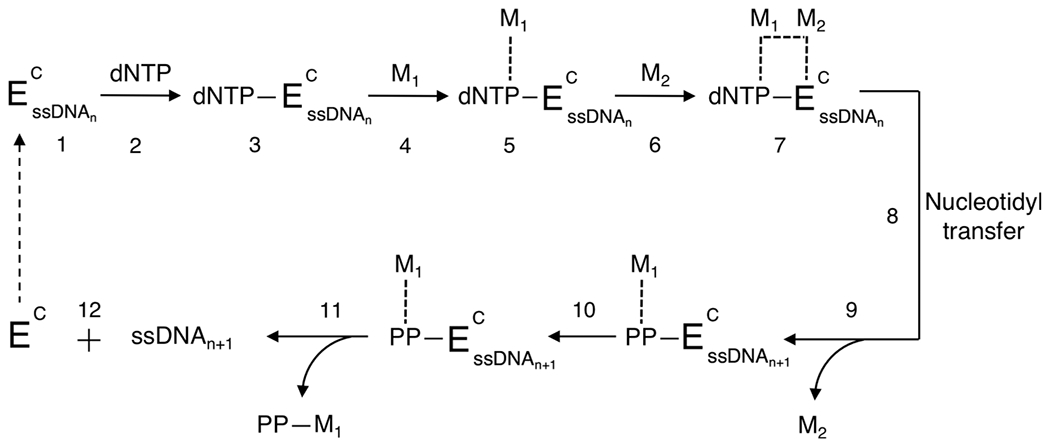
There is a growing demand for sustainable methods in research and development, where instead of hazardous chemicals, an aqueous medium is chosen to perform biological reactions. In this Perspective, we examine the history and current methodology of using enzymes to generate artificial single-stranded DNA. By using traditional solid-phase phosphoramidite chemistry as a metric, we also explore criteria for the method of template-independent enzymatic oligonucleotide synthesis (TiEOS). As its key component, we delve into the biology of one of the most enigmatic enzymes, terminal deoxynucleotidyl transferase (TdT). As TdT is found to exponentially increase antigen receptor diversity in the vertebrate immune system by adding nucleotides in a template-free manner, researchers have exploited this function as an alternative to the phosphoramidite synthesis method. Though TdT is currently the preferred enzyme for TiEOS, its random nucleotide incorporation presents a barrier in synthesis automation. Taking a closer look at the TiEOS cycle, particularly the coupling step, we find it is comprised of additions > n+1 and deletions. By tapping into the physical and biochemical properties of TdT, we strive to further elucidate its mercurial behavior and offer ways to better optimize TiEOS for production-grade oligonucleotide synthesis.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
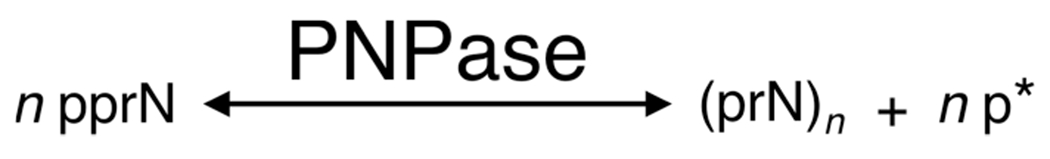
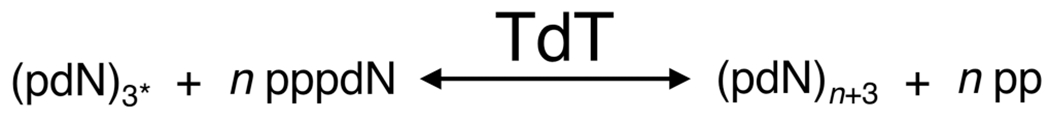

References
-
- Watson JD, and Crick FH (1953) Molecular structure of nucleic acids; a structure for deoxyribose nucleic acid. Nature 171, 737–738. - PubMed
-
- Lehman IR, Bessman MJ, Simms ES, and Kornberg A (1958) Enzymatic synthesis of deoxyribonucleic acid. I. Preparation of substrates and partial purification of an enzyme from Escherichia coli. J. Biol. Chem 233, 163–170. - PubMed
-
- Khorana HG (1968) Synthetic nucleic acids and the genetic code. JAMA, J. Am. Med. Assoc 206, 1978–1982. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
