

The Lung Microbiota of Healthy Mice Are Highly Variable, Cluster by Environment, and Reflect Variation in Baseline Lung Innate Immunity
- PMID: 29533677
- PMCID: PMC6118022
- DOI: 10.1164/rccm.201711-2180OC
The Lung Microbiota of Healthy Mice Are Highly Variable, Cluster by Environment, and Reflect Variation in Baseline Lung Innate Immunity
Abstract
Rationale: The "gut-lung axis" is commonly invoked to explain the microbiome's influence on lung inflammation. Yet the lungs harbor their own microbiome, which is altered in respiratory disease. The relative influence of gut and lung bacteria on lung inflammation is unknown.
Objectives: To determine whether baseline lung immune tone reflects local (lung-lung) or remote (gut-lung) microbe-host interactions.
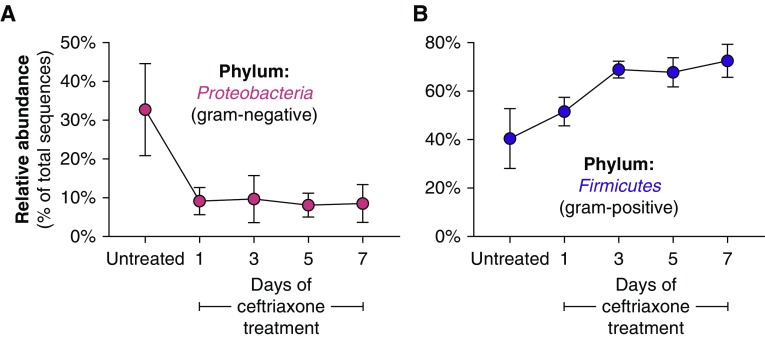
Methods: We compared lung, tongue, and cecal bacteria in 40 healthy, genetically identical, 10-week-old mice, using 16S ribosomal RNA gene quantification and sequencing. We measured inflammatory cytokines, using a multiplex assay of homogenized lung tissue. We compared lung bacteria in healthy mice treated with varied durations of systemic antibiotics.
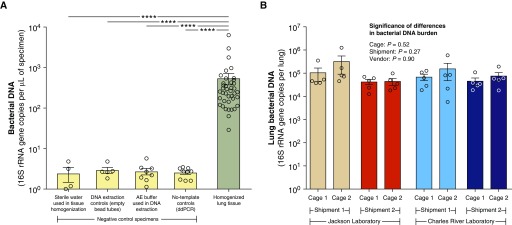
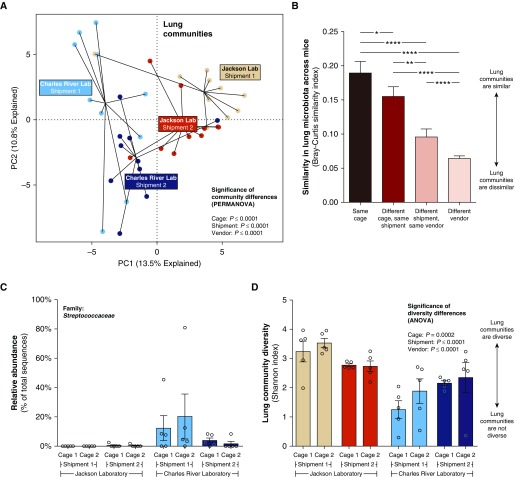
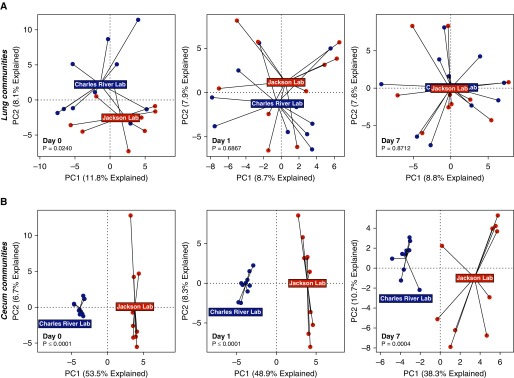
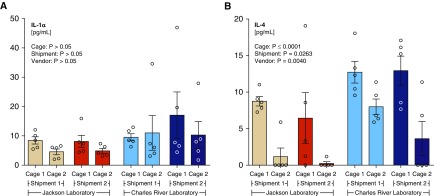
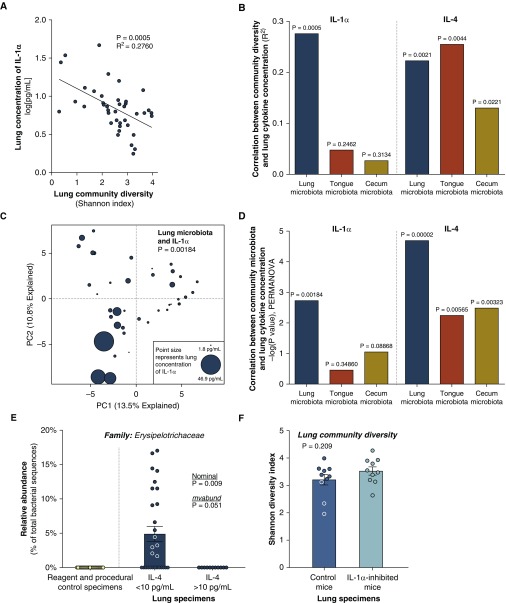
Measurements and main results: Lung bacterial communities are highly variable among mice, cluster strongly by cage, shipment, and vendor, and are altered by antibiotics in a microbiologically predictable manner. Baseline lung concentrations of two key inflammatory cytokines (IL-1α and IL-4) are correlated with the diversity and community composition of lung bacterial communities. Lung concentrations of these inflammatory cytokines correlate more strongly with variation in lung bacterial communities than with that of the gut or mouth.
Conclusions: In the lungs of healthy mice, baseline innate immune tone more strongly reflects local (lung-lung) microbe-host interactions than remote (gut-lung) microbe-host interactions. Our results independently confirm the existence and immunologic significance of the murine lung microbiome, even in health. Variation in lung microbiota is likely an important, underappreciated source of experimental and clinical variability. The lung microbiome is an unexplored therapeutic target for the prevention and treatment of inflammatory lung disease.
Keywords: 16S; host–microbiome interactions; innate immunity; lung microbiome.
Figures
Comment in
-
Of Mice and Men . . . and Microbes: Conclusions and Cautions from a Murine Study of the Lung Microbiome and Microbiome-Immune Interactions.Am J Respir Crit Care Med. 2018 Aug 15;198(4):419-422. doi: 10.1164/rccm.201803-0586ED. Am J Respir Crit Care Med. 2018. PMID: 29995432 Free PMC article. No abstract available.
References
-
- Ege MJ, Mayer M, Normand AC, Genuneit J, Cookson WO, Braun-Fahrländer C, et al. GABRIELA Transregio 22 Study Group. Exposure to environmental microorganisms and childhood asthma. N Engl J Med. 2011;364:701–709. - PubMed
-
- Weiss B, Bujanover Y, Yahav Y, Vilozni D, Fireman E, Efrati O. Probiotic supplementation affects pulmonary exacerbations in patients with cystic fibrosis: a pilot study. Pediatr Pulmonol. 2010;45:536–540. - PubMed
-
- Bruzzese E, Raia V, Spagnuolo MI, Volpicelli M, De Marco G, Maiuri L, et al. Effect of Lactobacillus GG supplementation on pulmonary exacerbations in patients with cystic fibrosis: a pilot study. Clin Nutr. 2007;26:322–328. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
