

Modulation of Inflammatory Arthritis in Mice by Gut Microbiota Through Mucosal Inflammation and Autoantibody Generation
- PMID: 29534332
- PMCID: PMC6105374
- DOI: 10.1002/art.40490
Modulation of Inflammatory Arthritis in Mice by Gut Microbiota Through Mucosal Inflammation and Autoantibody Generation
Abstract
Objective: Observations of microbial dysbiosis in patients with rheumatoid arthritis (RA) have raised interest in studying microbial-mucosal interactions as a potential trigger of RA. Using the murine collagen-induced arthritis (CIA) model, we undertook this study to test our hypothesis that microbiota modulate immune responses leading to autoimmune arthritis.
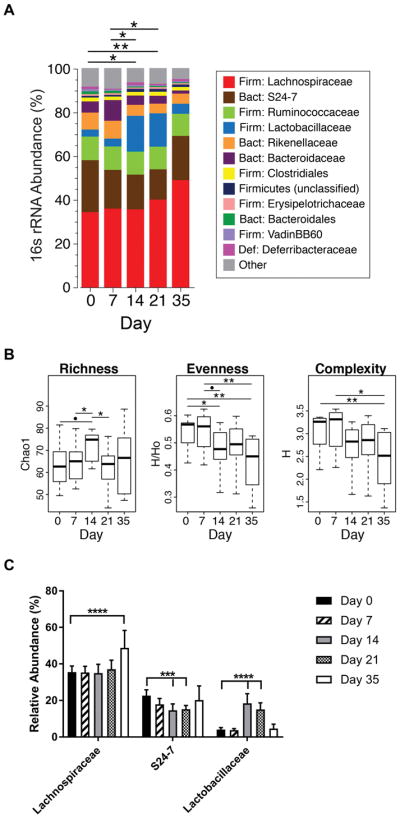
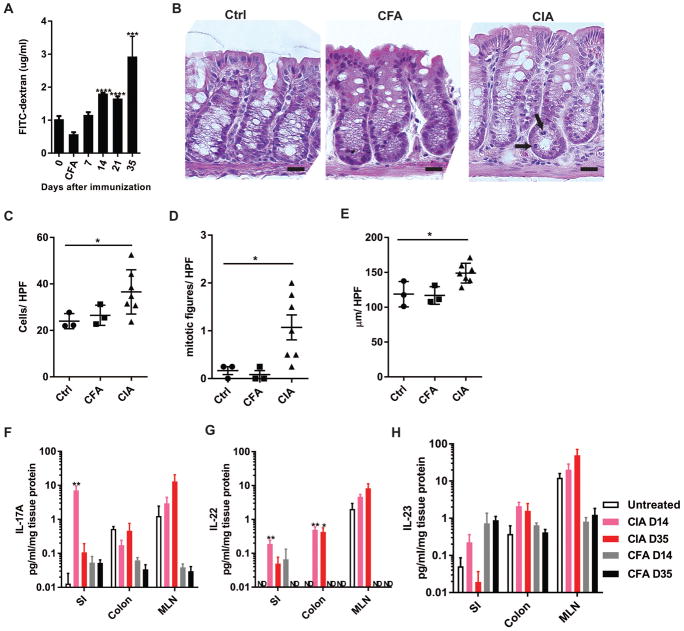
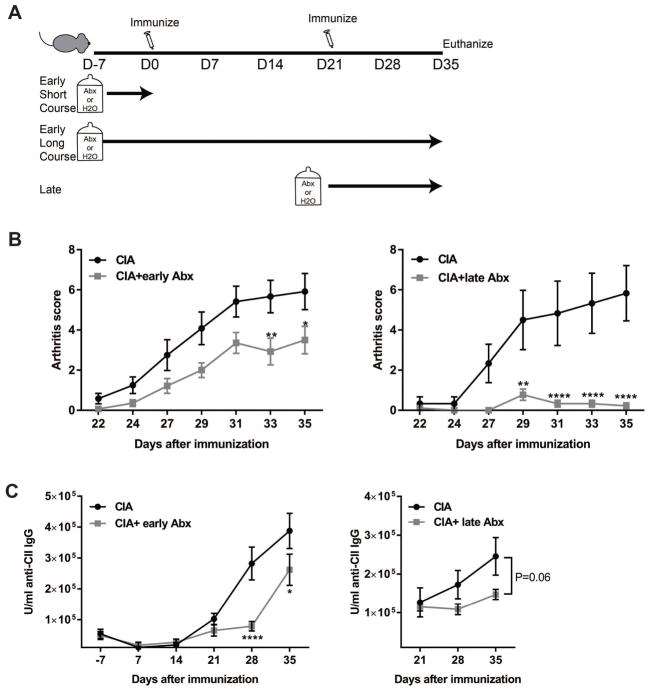
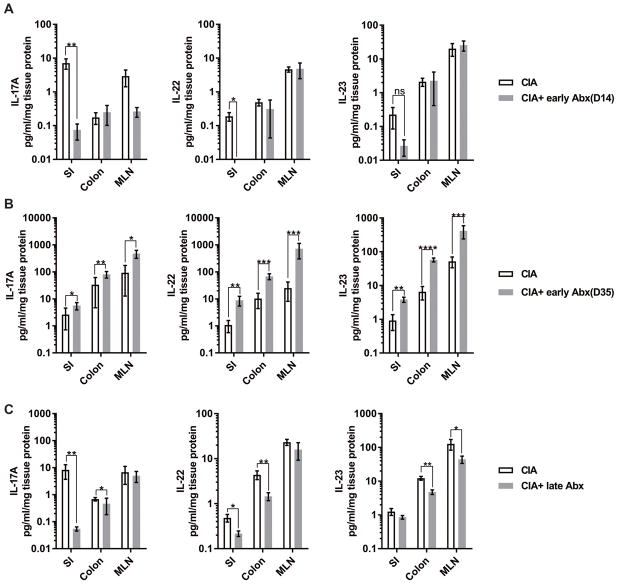
Methods: CIA was induced by immunization of mice with type II collagen (CII) in adjuvant on days 0 and 21, with arthritis appearing on days 23 and 24. Intestinal microbiota were profiled by 16S ribosomal RNA sequencing every 7 days during the course of CIA, and intestinal mucosal changes were evaluated on days 14 and 35. Then, microbiota were depleted either early (7 days before immunization) or late (day 21 after immunization) by administration of broad-spectrum antibiotics. Disease severity, autoantibody and systemic cytokine production, and intestinal mucosal responses were monitored in the setting of microbial reduction.
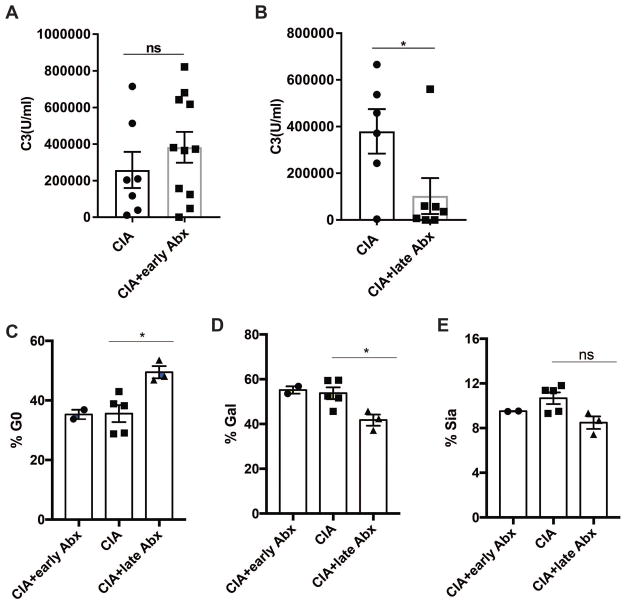
Results: Significant dysbiosis and mucosal inflammation occurred early in CIA, prior to visible arthritis, and continued to evolve during the course of disease. Depletion of the microbiota prior to the induction of CIA resulted in an ~40% reduction in disease severity and in significantly reduced levels of serum inflammatory cytokines and anti-CII antibodies. In intestinal tissue, production of interleukin-17A (IL-17A) and IL-22 was delayed. Unexpectedly, microbial depletion during the late phase of CIA resulted in a >50% decrease in disease severity. Anti-CII antibodies were mildly reduced but were significantly impaired in their ability to activate complement, likely due to altered glycosylation profiles.
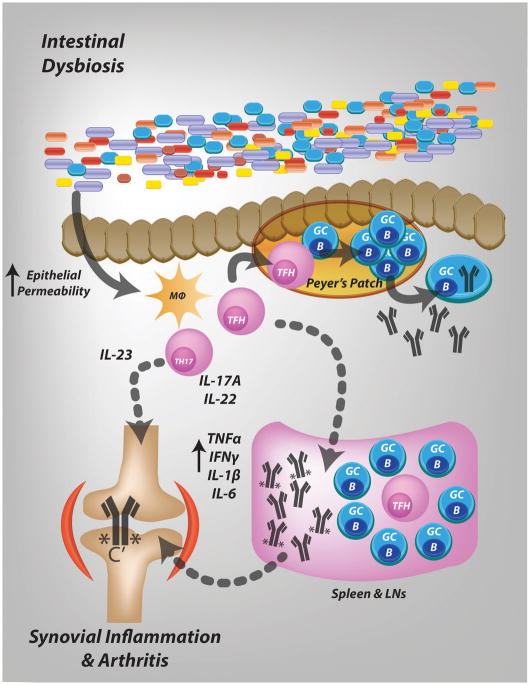
Conclusion: These data support a model in which intestinal dysbiosis triggers mucosal immune responses that stimulate T and B cells that are key for the development of inflammatory arthritis.
© 2018, American College of Rheumatology.
Conflict of interest statement
Figures
References
-
- Tobon GJ, Youinou P, Saraux A. The environment, geo-epidemiology, and autoimmune disease: Rheumatoid arthritis. J Autoimmun. 2010;35(1):10–14. - PubMed
-
- Padyukov L, Silva C, Stolt P, Alfredsson L, Klareskog L. A gene-environment interaction between smoking and shared epitope genes in HLA-DR provides a high risk of seropositive rheumatoid arthritis. Arthritis Rheum. 2004;50(10):3085–3092. - PubMed
-
- van Gaalen FA, Linn-Rasker SP, van Venrooij WJ, de Jong BA, Breedveld FC, Verweij CL, Toes RE, Huizinga TW. Autoantibodies to cyclic citrullinated peptides predict progression to rheumatoid arthritis in patients with undifferentiated arthritis: a prospective cohort study. Arthritis Rheum. 2004;50(3):709–715. - PubMed
-
- Shi J, Knevel R, Suwannalai P, van der Linden MP, Janssen GM, van Veelen PA, Levarht NE, van der Helm-van Mil AH, Cerami A, Huizinga TW, et al. Autoantibodies recognizing carbamylated proteins are present in sera of patients with rheumatoid arthritis and predict joint damage. Proc Natl Acad Sci U S A. 2011;108(42):17372–17377. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
