

Extension of chemotactic pseudopods by nonadherent human neutrophils does not require or cause calcium bursts
- PMID: 29535263
- PMCID: PMC7053518
- DOI: 10.1126/scisignal.aal4289
Extension of chemotactic pseudopods by nonadherent human neutrophils does not require or cause calcium bursts
Abstract
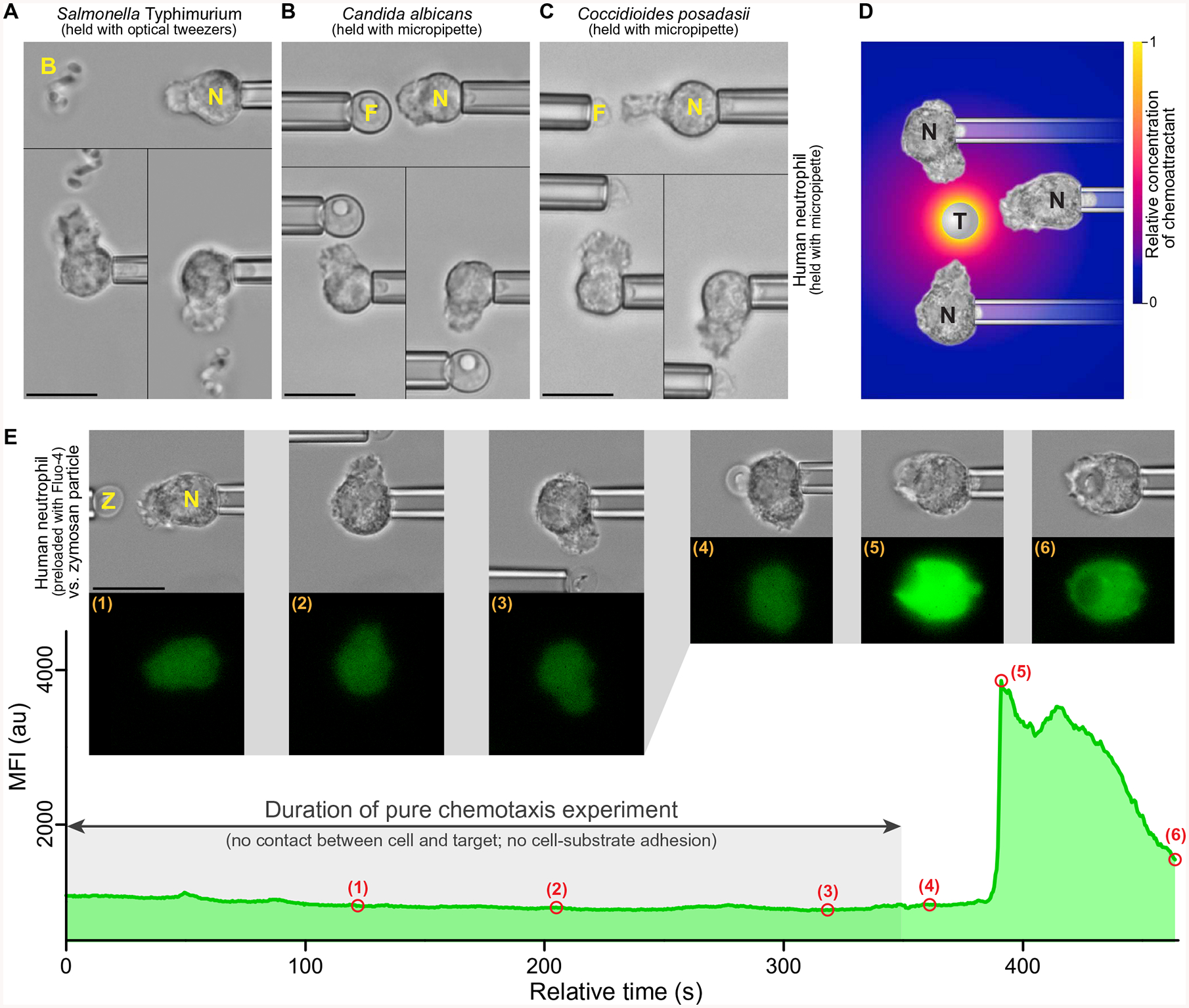
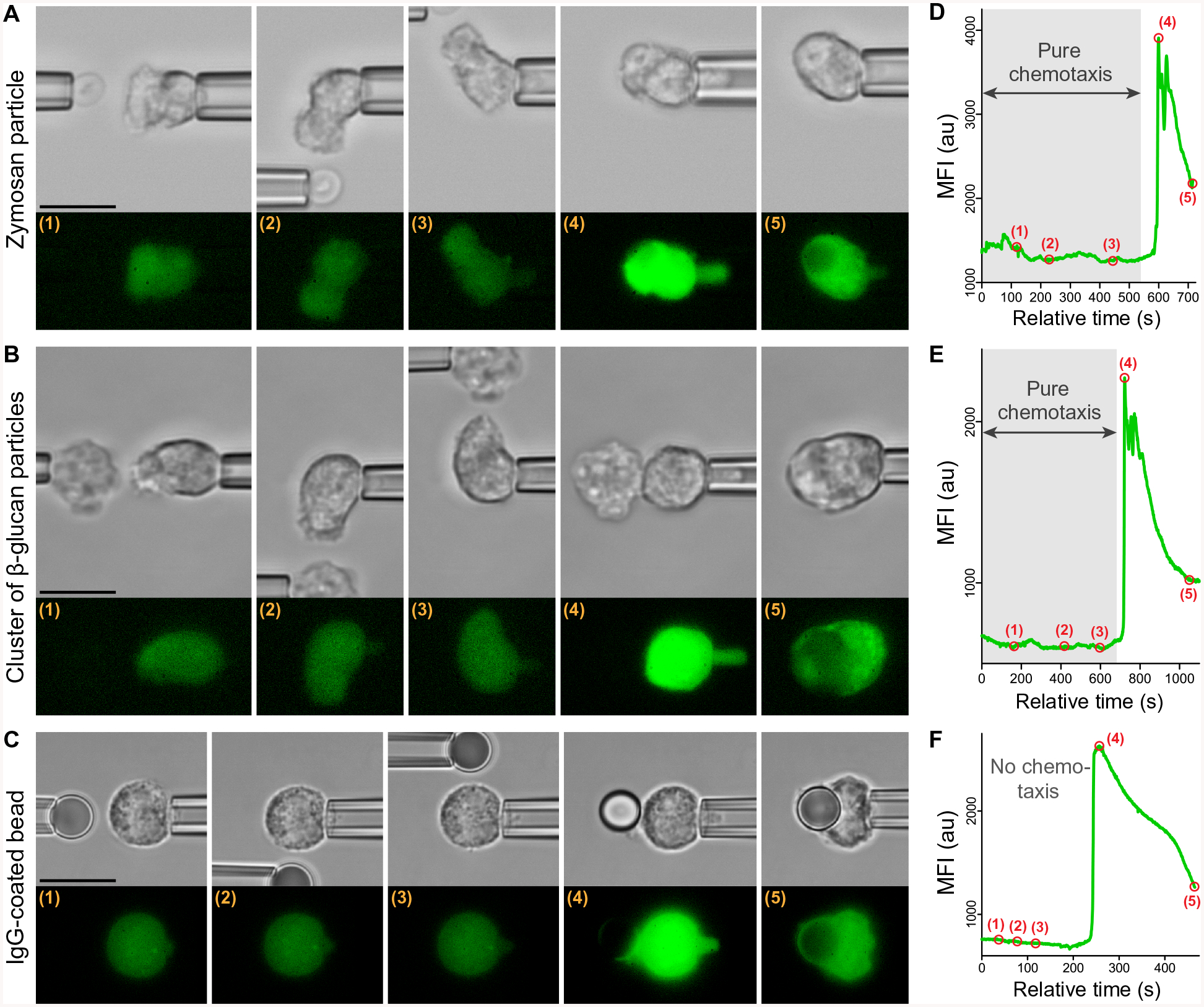
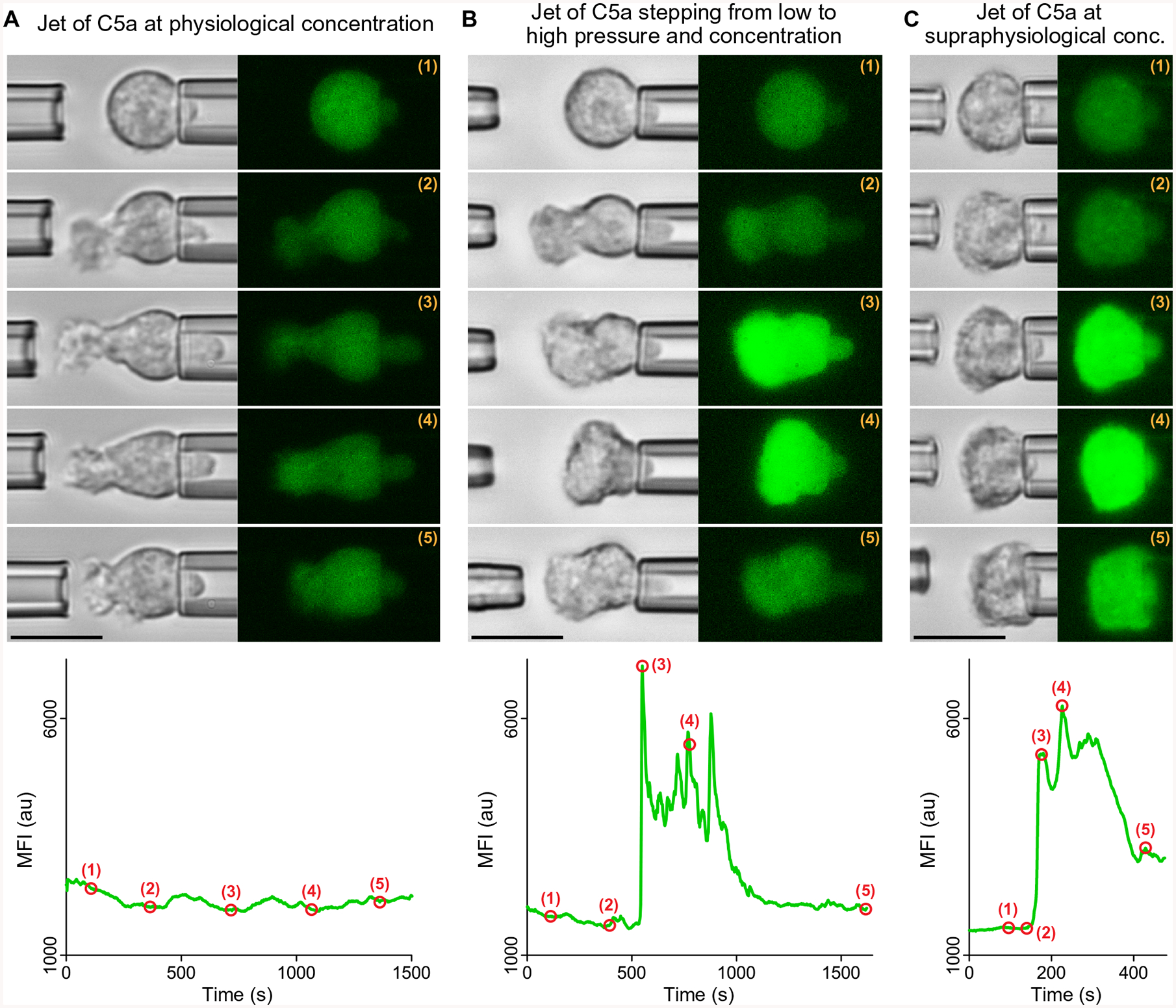
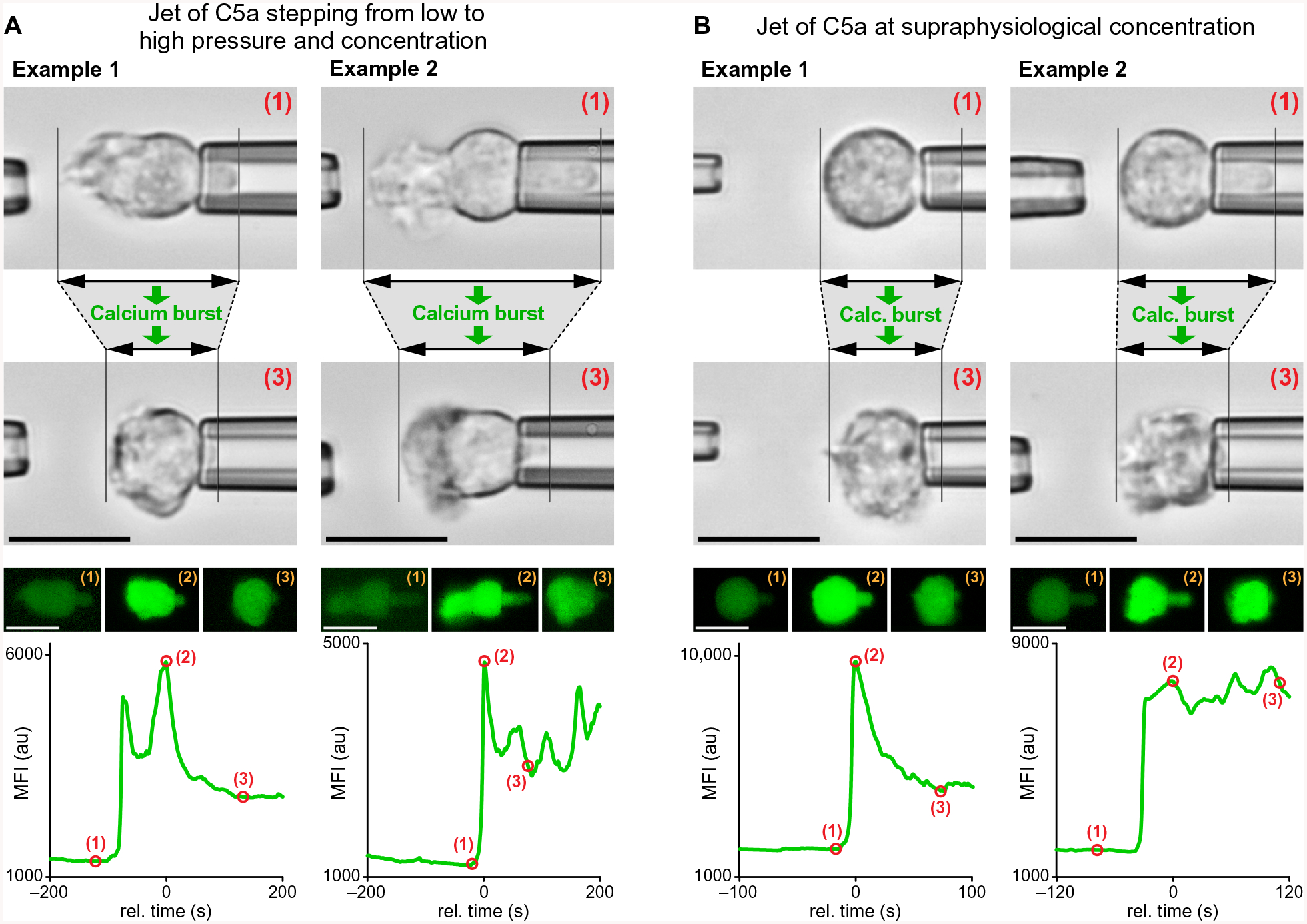
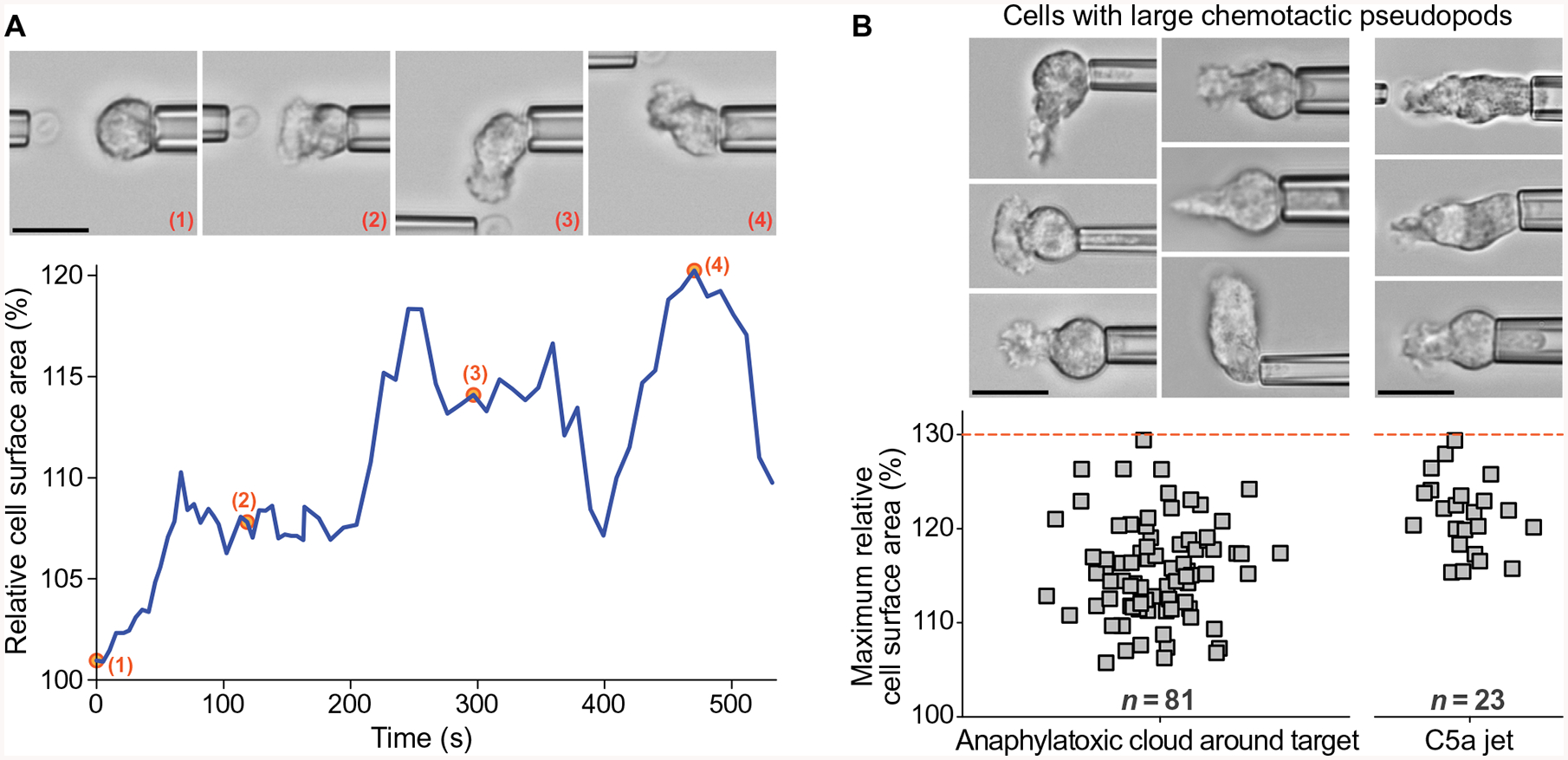
Global bursts in free intracellular calcium (Ca2+) are among the most conspicuous signaling events in immune cells. To test the common view that Ca2+ bursts mediate rearrangement of the actin cytoskeleton in response to the activation of G protein-coupled receptors, we combined single-cell manipulation with fluorescence imaging and monitored the Ca2+ concentration in individual human neutrophils during complement-mediated chemotaxis. By decoupling purely chemotactic pseudopod formation from cell-substrate adhesion, we showed that physiological concentrations of anaphylatoxins, such as C5a, induced nonadherent human neutrophils to form chemotactic pseudopods but did not elicit Ca2+ bursts. By contrast, pathological or supraphysiological concentrations of C5a often triggered Ca2+ bursts, but pseudopod protrusion stalled or reversed in such cases, effectively halting chemotaxis, similar to sepsis-associated neutrophil paralysis. The maximum increase in cell surface area during pseudopod extension in pure chemotaxis was much smaller-by a factor of 8-than the known capacity of adherent human neutrophils to expand their surface. Because the measured rise in cortical tension was not sufficient to account for this difference, we attribute the limited deformability to a reduced ability of the cytoskeleton to generate protrusive force in the absence of cell adhesion. Thus, we hypothesize that Ca2+ bursts in neutrophils control a mechanistic switch between two distinct modes of cytoskeletal organization and dynamics. A key element of this switch appears to be the expedient coordination of adhesion-dependent lock or release events of cytoskeletal membrane anchors.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
Similar articles
-
Complement C5a-Induced Changes in Neutrophil Morphology During Inflammation.Scand J Immunol. 2017 Sep;86(3):143-155. doi: 10.1111/sji.12580. Scand J Immunol. 2017. PMID: 28671713 Free PMC article.
-
Controlled pseudopod extension of human neutrophils stimulated with different chemoattractants.Biophys J. 2004 Jul;87(1):688-95. doi: 10.1529/biophysj.103.036699. Biophys J. 2004. PMID: 15240502 Free PMC article.
-
Anaphylatoxin signaling in human neutrophils. A key role for sphingosine kinase.J Biol Chem. 2004 Oct 22;279(43):44802-11. doi: 10.1074/jbc.M403977200. Epub 2004 Aug 9. J Biol Chem. 2004. PMID: 15302883
-
The role of the complement anaphylatoxins in the recruitment of eosinophils.Int Immunopharmacol. 2007 Dec 20;7(14):1909-23. doi: 10.1016/j.intimp.2007.07.006. Epub 2007 Aug 6. Int Immunopharmacol. 2007. PMID: 18039528 Review.
-
Role of C5a-C5aR interaction in sepsis.Shock. 2004 Jan;21(1):1-7. doi: 10.1097/01.shk.0000105502.75189.5e. Shock. 2004. PMID: 14676676 Review.
Cited by
-
Micropipette-based biomechanical nanotools on living cells.Eur Biophys J. 2022 Mar;51(2):119-133. doi: 10.1007/s00249-021-01587-5. Epub 2022 Feb 16. Eur Biophys J. 2022. PMID: 35171346 Free PMC article. Review.
-
Mechanistic Understanding of Single-Cell Behavior is Essential for Transformative Advances in Biomedicine.Yale J Biol Med. 2018 Sep 21;91(3):279-289. eCollection 2018 Sep. Yale J Biol Med. 2018. PMID: 30258315 Free PMC article. Review.
-
TRPM2 ion channels steer neutrophils towards a source of hydrogen peroxide.Sci Rep. 2021 Apr 29;11(1):9339. doi: 10.1038/s41598-021-88224-5. Sci Rep. 2021. PMID: 33927223 Free PMC article.
-
Localisation of Intracellular Signals and Responses during Phagocytosis.Int J Mol Sci. 2023 Feb 1;24(3):2825. doi: 10.3390/ijms24032825. Int J Mol Sci. 2023. PMID: 36769146 Free PMC article. Review.
-
Biophysical nanotools for single-molecule dynamics.Biophys Rev. 2018 Oct;10(5):1349-1357. doi: 10.1007/s12551-018-0447-y. Epub 2018 Aug 18. Biophys Rev. 2018. PMID: 30121743 Free PMC article. Review.
References
-
- White JR, Naccache PH, Molski TFP, Borgeat P, Sha’afi RI, Direct demonstration of increased intracellular concentration of free calcium in rabbit and human neutrophils following stimulation by chemotactic factor. Biochem. Biophys. Res. Commun 113, 44–50 (1983). - PubMed
-
- Lew DP, Receptor signalling and intracellular calcium in neutrophil activation. Eur. J. Clin. Invest 19, 338–346 (1989). - PubMed
-
- Norgauer J, Dobos G, Kownatzki E, Dahinden C, Burger R, Kupper R, Gierschik P, Complement fragment C3a stimulates Ca2+ influx in neutrophils via a pertussis-toxin-sensitive G protein. Eur. J. Biochem 217, 289–294 (1993). - PubMed
-
- Schorr W, Swandulla D, Zeilhofer HU, Mechanisms of IL-8-induced Ca2+ signaling in human neutrophil granulocytes. Eur. J. Immunol 29, 897–904 (1999). - PubMed
-
- Ibrahim FB, Pang SJ, Melendez AJ, Anaphylatoxin signaling in human neutrophils. A key role for sphingosine kinase. J. Biol. Chem 279, 44802–44811 (2004). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
