

Musashi and Plasticity of Xenopus and Axolotl Spinal Cord Ependymal Cells
- PMID: 29535610
- PMCID: PMC5835034
- DOI: 10.3389/fncel.2018.00045
Musashi and Plasticity of Xenopus and Axolotl Spinal Cord Ependymal Cells
Abstract
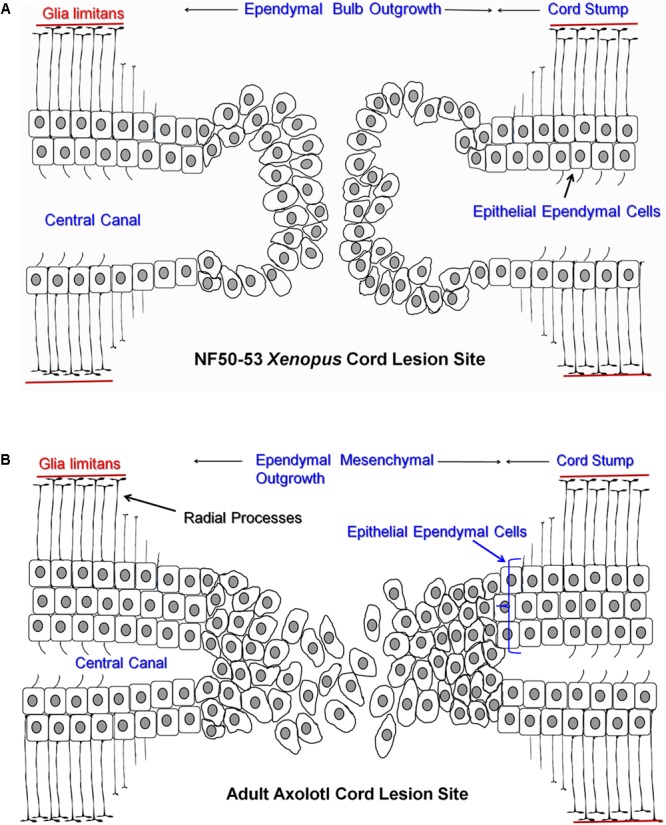
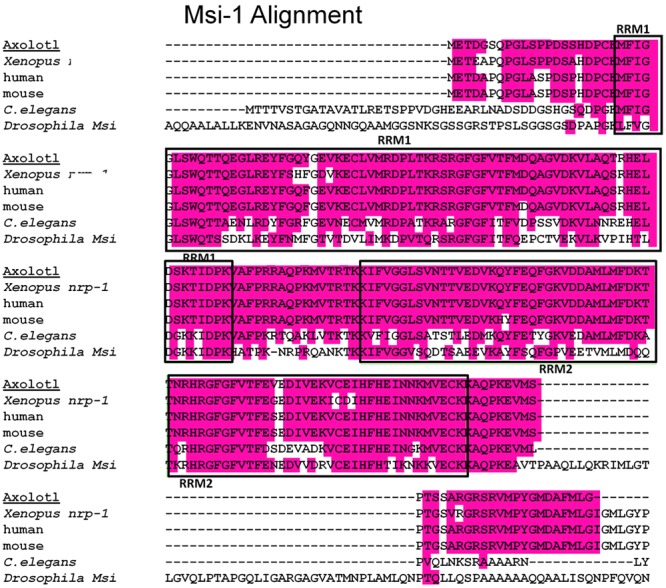
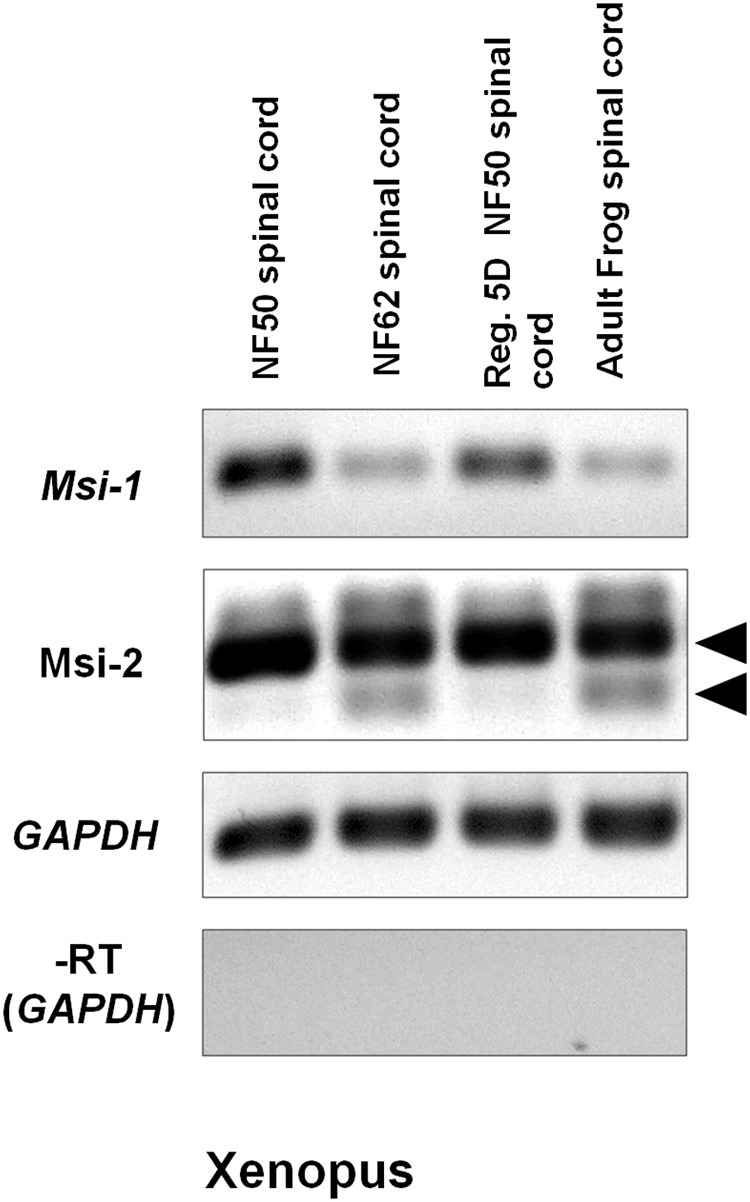
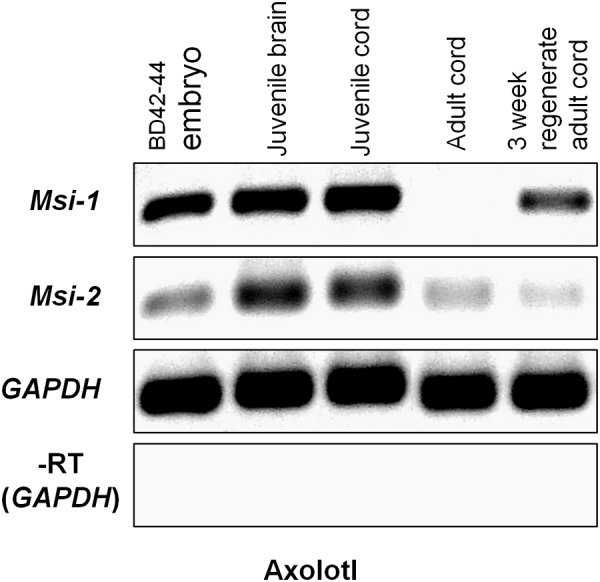
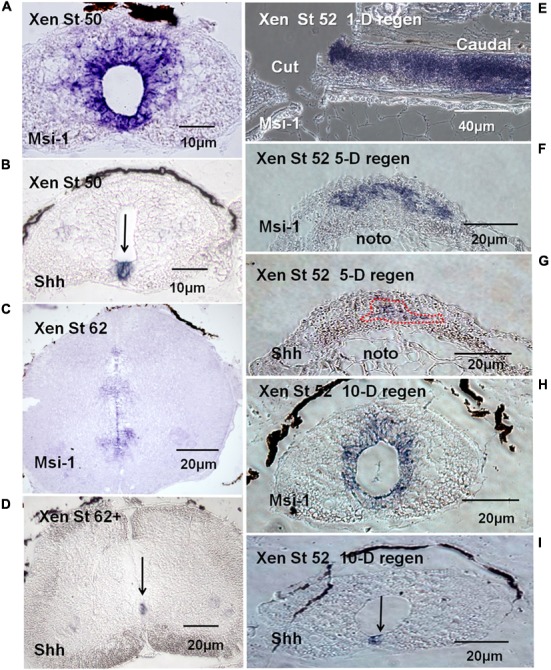
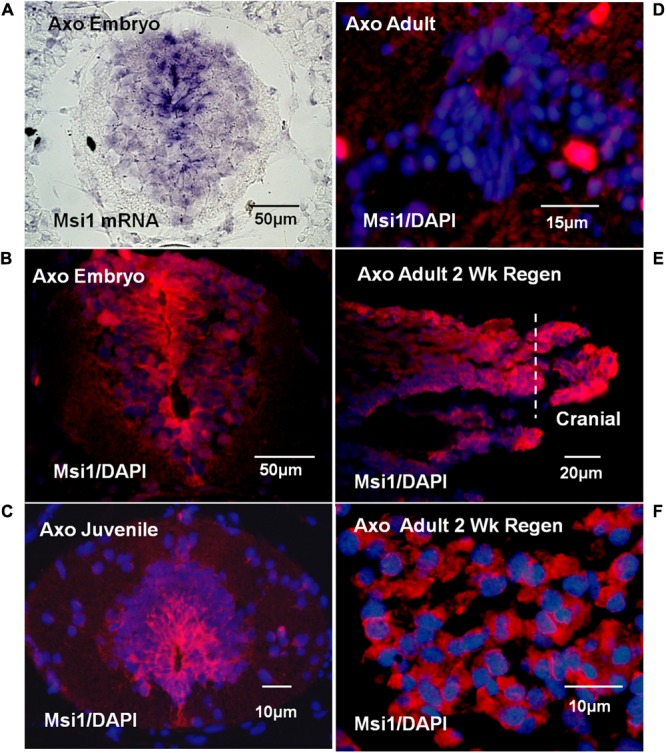
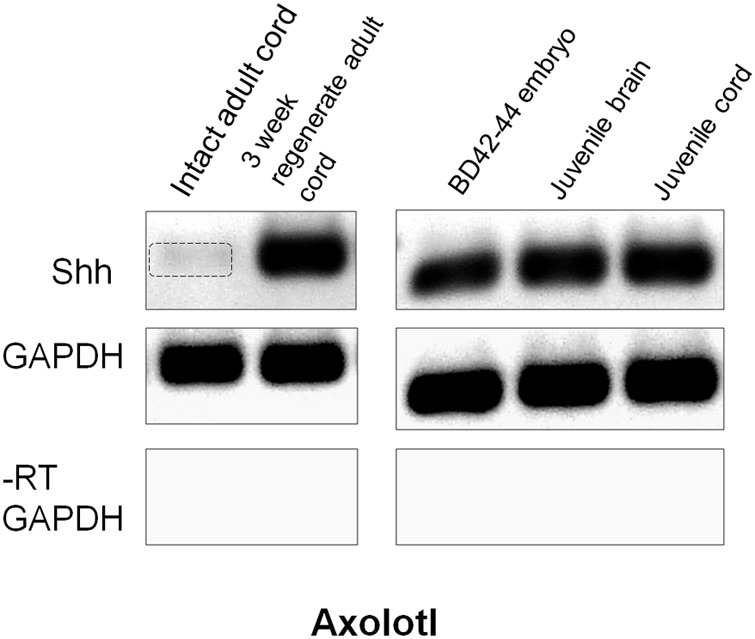
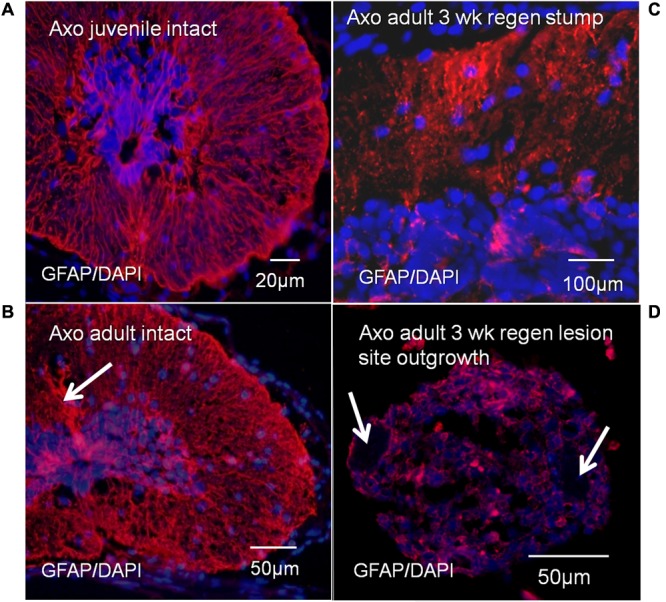
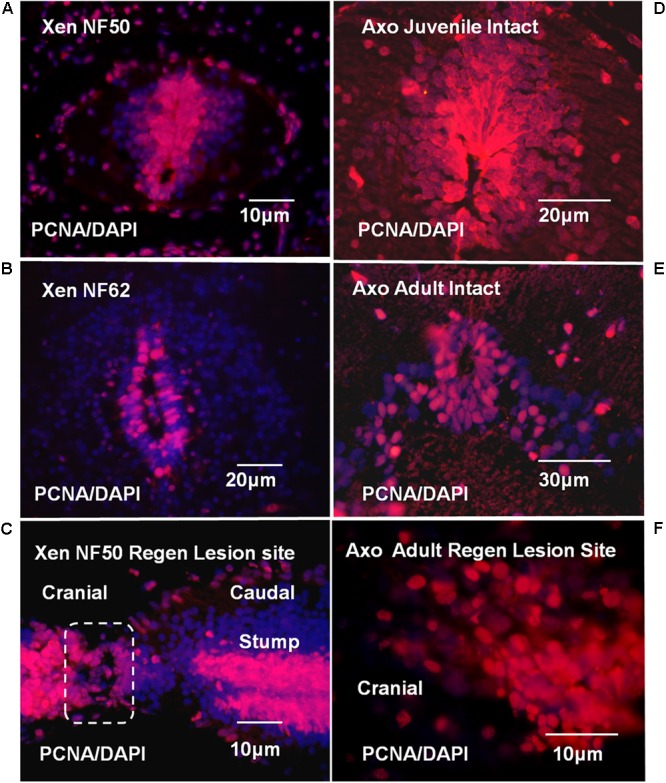
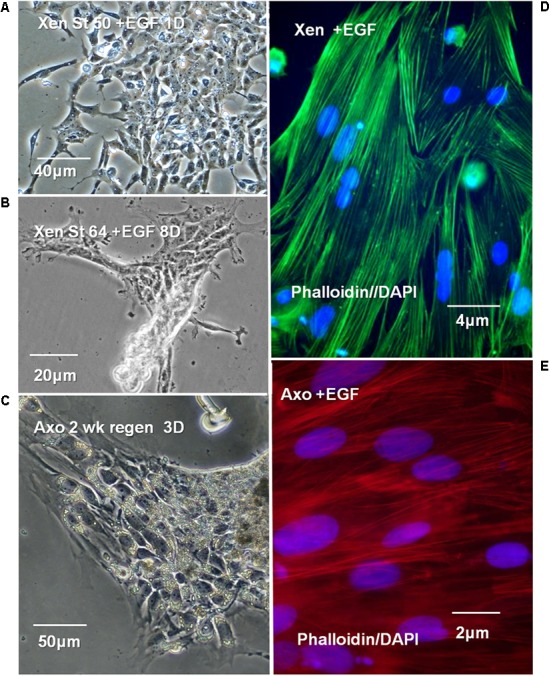
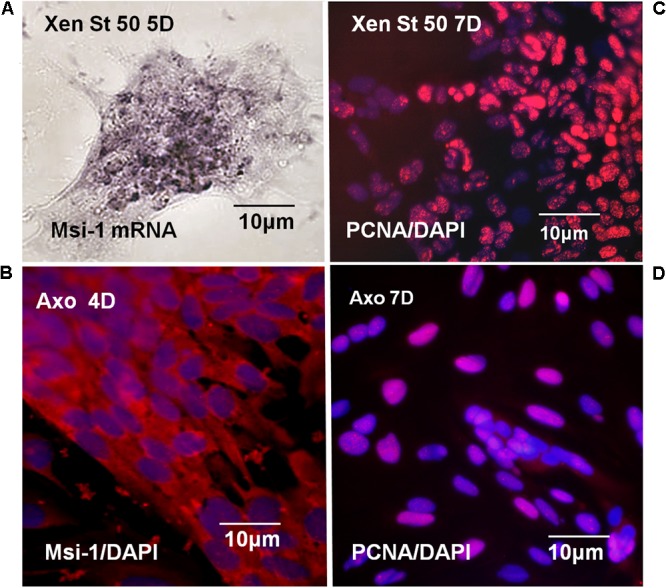
The differentiated state of spinal cord ependymal cells in regeneration-competent amphibians varies between a constitutively active state in what is essentially a developing organism, the tadpole of the frog Xenopus laevis, and a quiescent, activatable state in a slowly growing adult salamander Ambystoma mexicanum, the Axolotl. Ependymal cells are epithelial in intact spinal cord of all vertebrates. After transection, body region ependymal epithelium in both Xenopus and the Axolotl disorganizes for regenerative outgrowth (gap replacement). Injury-reactive ependymal cells serve as a stem/progenitor cell population in regeneration and reconstruct the central canal. Expression patterns of mRNA and protein for the stem/progenitor cell-maintenance Notch signaling pathway mRNA-binding protein Musashi (msi) change with life stage and regeneration competence. Msi-1 is missing (immunohistochemistry), or at very low levels (polymerase chain reaction, PCR), in both intact regeneration-competent adult Axolotl cord and intact non-regeneration-competent Xenopus tadpole (Nieuwkoop and Faber stage 62+, NF 62+). The critical correlation for successful regeneration is msi-1 expression/upregulation after injury in the ependymal outgrowth and stump-region ependymal cells. msi-1 and msi-2 isoforms were cloned for the Axolotl as well as previously unknown isoforms of Xenopus msi-2. Intact Xenopus spinal cord ependymal cells show a loss of msi-1 expression between regeneration-competent (NF 50-53) and non-regenerating stages (NF 62+) and in post-metamorphosis froglets, while msi-2 displays a lower molecular weight isoform in non-regenerating cord. In the Axolotl, embryos and juveniles maintain Msi-1 expression in the intact cord. In the adult Axolotl, Msi-1 is absent, but upregulates after injury. Msi-2 levels are more variable among Axolotl life stages: rising between late tailbud embryos and juveniles and decreasing in adult cord. Cultures of regeneration-competent Xenopus tadpole cord and injury-responsive adult Axolotl cord ependymal cells showed an identical growth factor response. Epidermal growth factor (EGF) maintains mesenchymal outgrowth in vitro, the cells are proliferative and maintain msi-1 expression. Non-regeneration competent Xenopus ependymal cells, NF 62+, failed to attach or grow well in EGF+ medium. Ependymal Msi-1 expression in vivo and in vitro is a strong indicator of regeneration competence in the amphibian spinal cord.
Keywords: Axolotl regeneration; Xenopus regeneration; ependymal cells; musashi-1; musashi-2; spinal cord regeneration.
Figures


References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous