

Recurrent excitation between motoneurones propagates across segments and is purely glutamatergic
- PMID: 29538375
- PMCID: PMC5851534
- DOI: 10.1371/journal.pbio.2003586
Recurrent excitation between motoneurones propagates across segments and is purely glutamatergic
Abstract
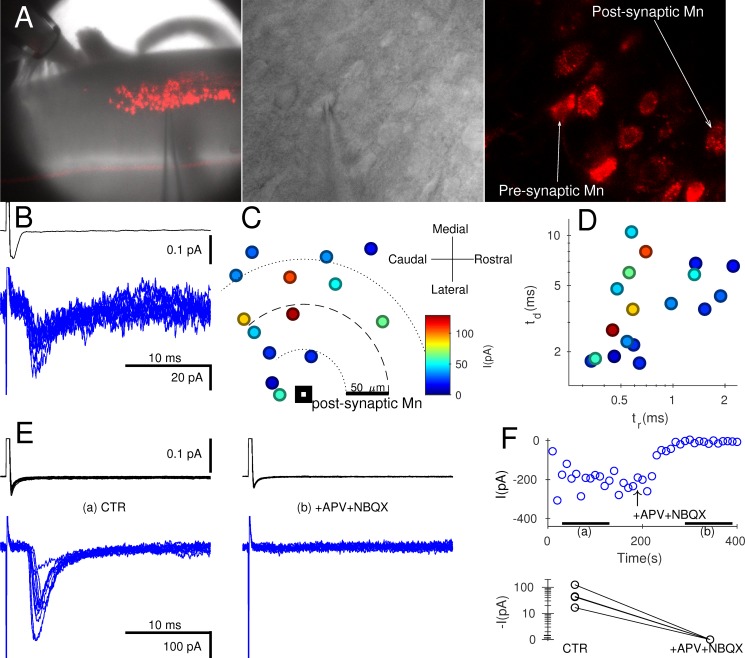
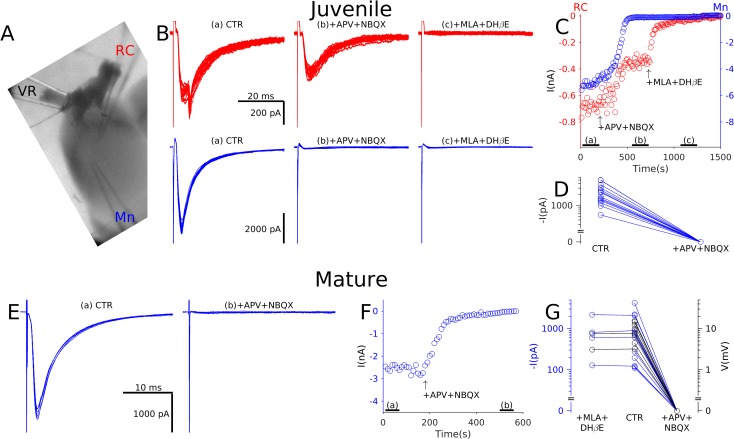
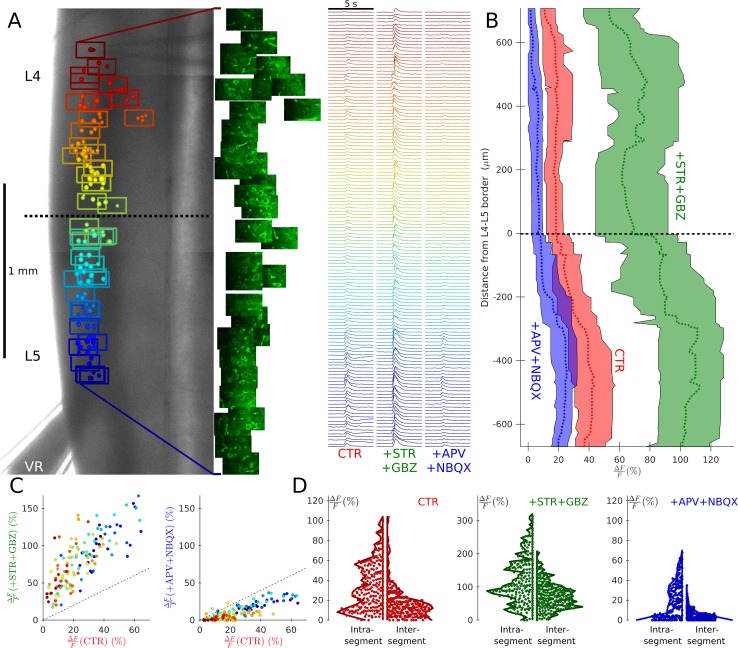
Spinal motoneurones (Mns) constitute the final output for the execution of motor tasks. In addition to innervating muscles, Mns project excitatory collateral connections to Renshaw cells (RCs) and other Mns, but the latter have received little attention. We show that Mns receive strong synaptic input from other Mns throughout development and into maturity, with fast-type Mns systematically receiving greater recurrent excitation than slow-type Mns. Optical recordings show that activation of Mns in one spinal segment can propagate to adjacent segments even in the presence of intact recurrent inhibition. While it is known that transmission at the neuromuscular junction is purely cholinergic and RCs are excited through both acetylcholine and glutamate receptors, here we show that neurotransmission between Mns is purely glutamatergic, indicating that synaptic transmission systems are differentiated at different postsynaptic targets of Mns.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


References
-
- Kiehn O. Decoding the organization of spinal circuits that control locomotion. Nat Rev Neurosci. 2016;17(4):224–38. doi: 10.1038/nrn.2016.9 - DOI - PMC - PubMed
-
- Cullheim S, Kellerth JO, Conradi S. Evidence for direct synaptic interconnections between cat spinal alpha-motoneurons via the recurrent axon collaterals: a morphological study using intracellular injection of horseradish peroxidase. Brain Res. 1977;132(1):1–10. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
