

A Flow Cytometry-Based Phenotypic Screen To Identify Novel Endocytic Factors in Saccharomyces cerevisiae
- PMID: 29540444
- PMCID: PMC5940143
- DOI: 10.1534/g3.118.200102
A Flow Cytometry-Based Phenotypic Screen To Identify Novel Endocytic Factors in Saccharomyces cerevisiae
Abstract
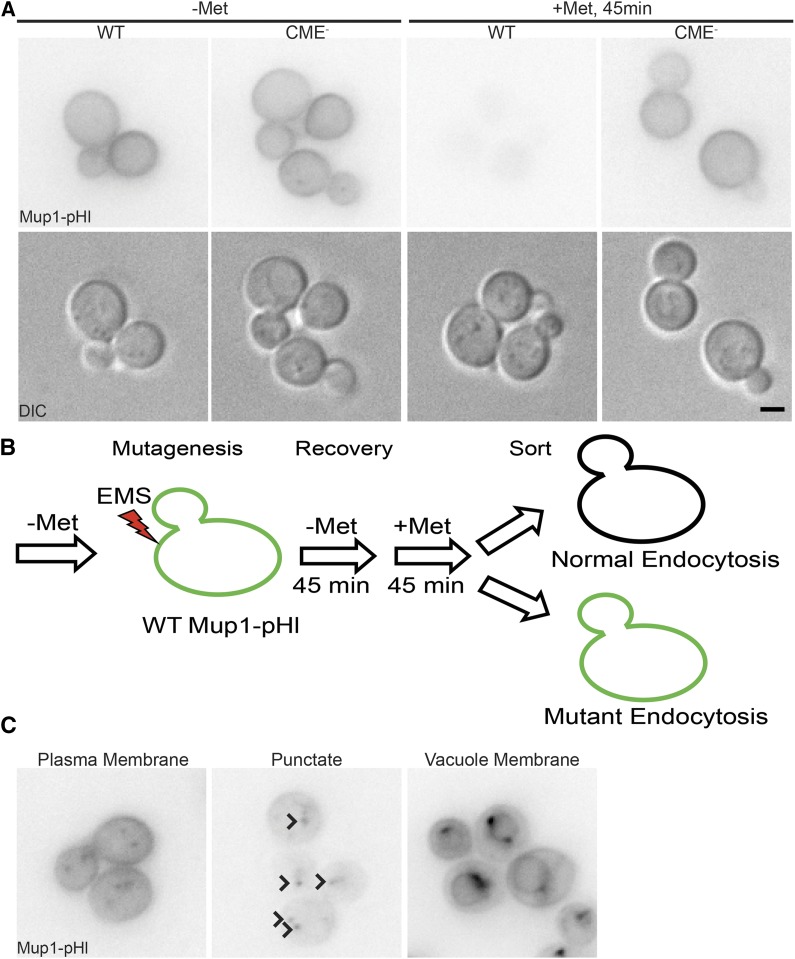
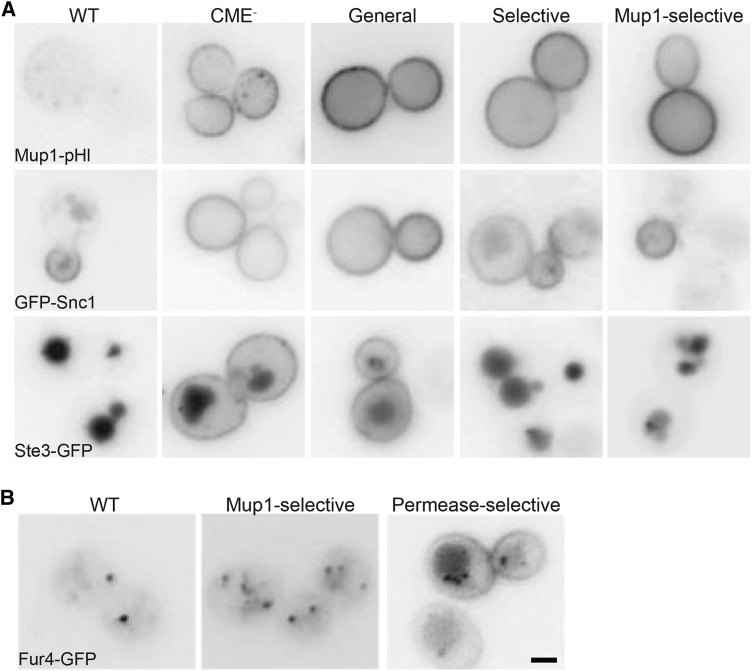
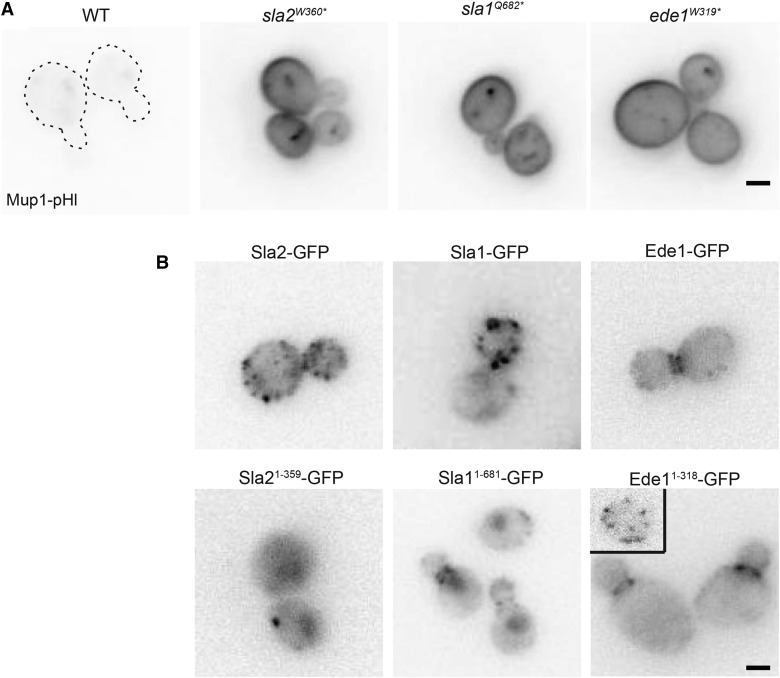
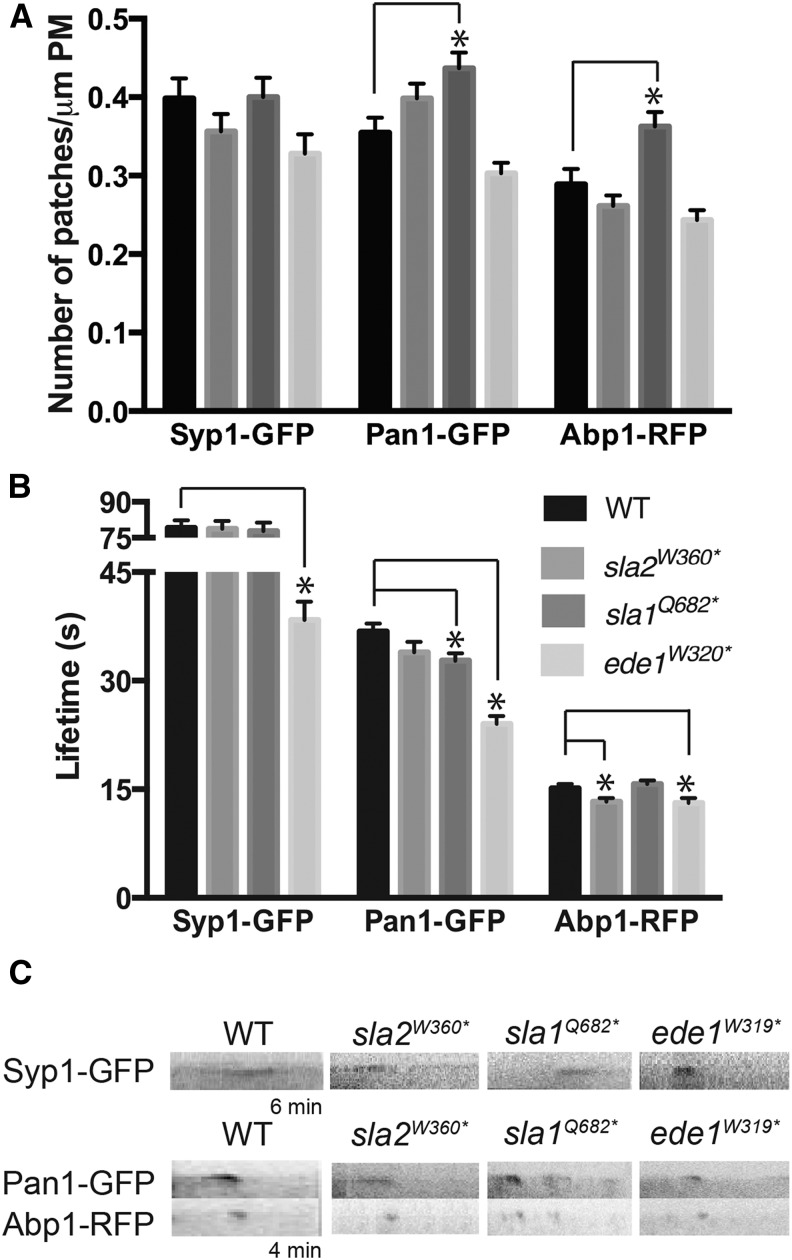
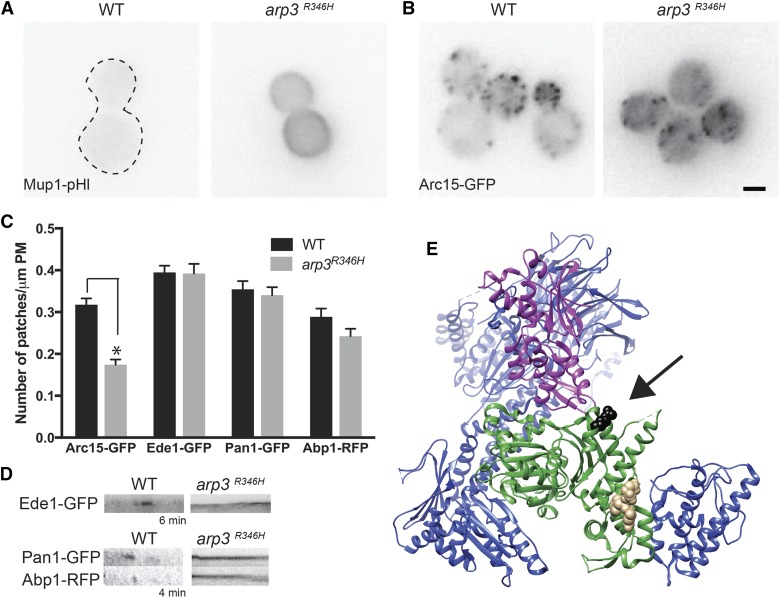
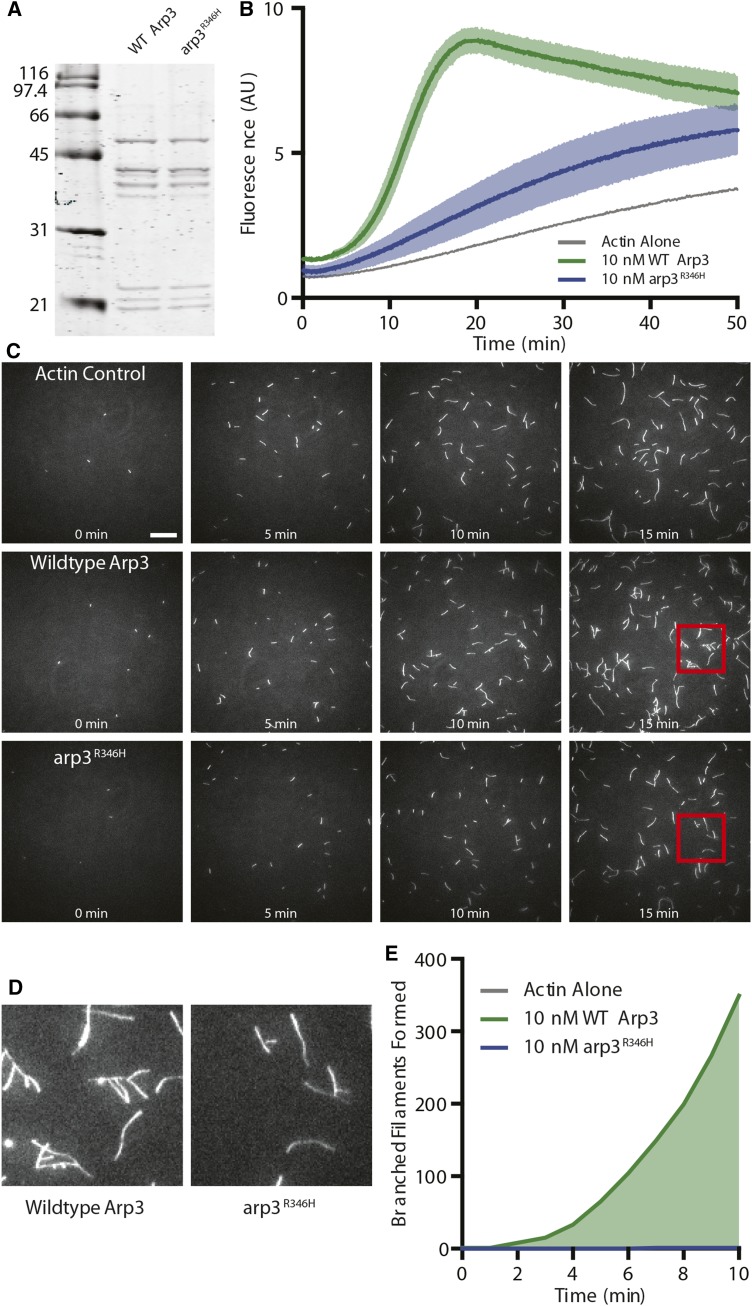
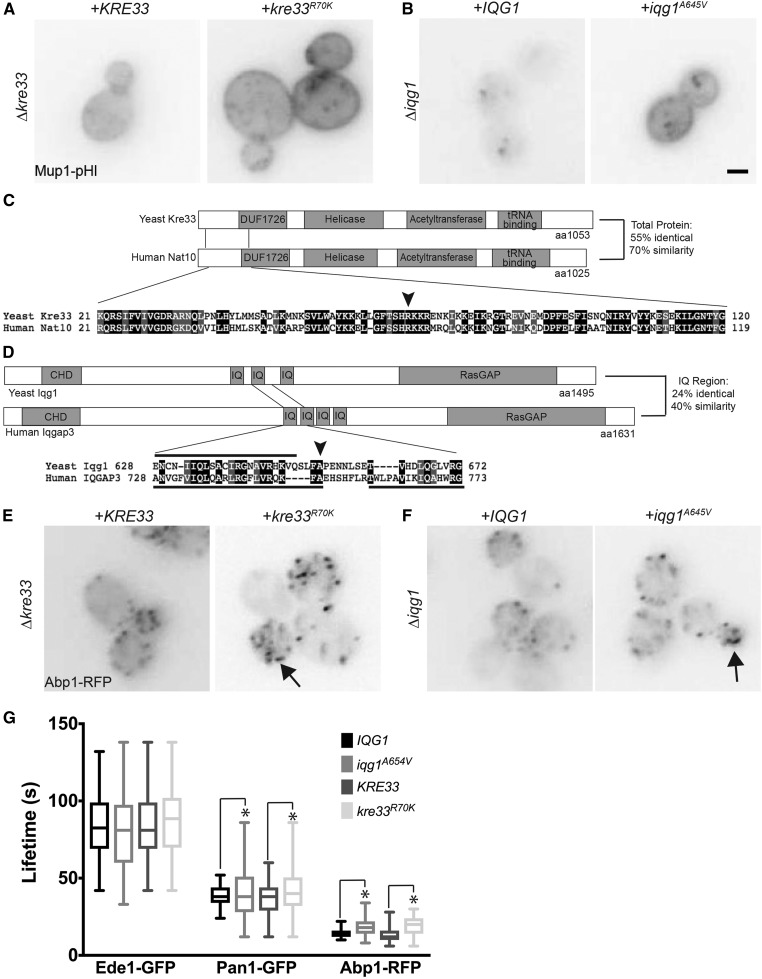
Endocytosis is a fundamental process for internalizing material from the plasma membrane, including many transmembrane proteins that are selectively internalized depending on environmental conditions. In most cells, the main route of entry is clathrin-mediated endocytosis (CME), a process that involves the coordinated activity of over 60 proteins; however, there are likely as-yet unidentified proteins involved in cargo selection and/or regulation of endocytosis. We performed a mutagenic screen to identify novel endocytic genes in Saccharomyces cerevisiae expressing the methionine permease Mup1 tagged with pHluorin (pHl), a pH-sensitive GFP variant whose fluorescence is quenched upon delivery to the acidic vacuole lumen. We used fluorescence-activated cell sorting to isolate mutagenized cells with elevated fluorescence, resulting from failure to traffic Mup1-pHl cargo to the vacuole, and further assessed subcellular localization of Mup1-pHl to characterize the endocytic defects in 256 mutants. A subset of mutant strains was classified as having general endocytic defects based on mislocalization of additional cargo proteins. Within this group, we identified mutations in four genes encoding proteins with known roles in endocytosis: the endocytic coat components SLA2, SLA1, and EDE1, and the ARP3 gene, whose product is involved in nucleating actin filaments to form branched networks. All four mutants demonstrated aberrant dynamics of the endocytic machinery at sites of CME; moreover, the arp3R346H mutation showed reduced actin nucleation activity in vitro Finally, whole genome sequencing of two general endocytic mutants identified mutations in conserved genes not previously implicated in endocytosis, KRE33 and IQG1, demonstrating that our screening approach can be used to identify new components involved in endocytosis.
Keywords: Arp2/3; actin; cargo sorting; clathrin-mediated endocytosis; forward genetic screen.
Copyright © 2018 Wrasman et al.
Figures
Similar articles
-
New Regulators of Clathrin-Mediated Endocytosis Identified in Saccharomyces cerevisiae by Systematic Quantitative Fluorescence Microscopy.Genetics. 2015 Nov;201(3):1061-70. doi: 10.1534/genetics.115.180729. Epub 2015 Sep 10. Genetics. 2015. PMID: 26362318 Free PMC article.
-
Sla1p serves as the targeting signal recognition factor for NPFX(1,2)D-mediated endocytosis.J Cell Biol. 2002 Apr 15;157(2):315-26. doi: 10.1083/jcb.200110027. Epub 2002 Apr 8. J Cell Biol. 2002. PMID: 11940605 Free PMC article.
-
ESCRT-dependent protein sorting is required for the viability of yeast clathrin-mediated endocytosis mutants.Traffic. 2020 Jun;21(6):430-450. doi: 10.1111/tra.12731. Traffic. 2020. PMID: 32255230 Free PMC article.
-
Function and regulation of Saccharomyces cerevisiae myosins-I in endocytic budding.Biochem Soc Trans. 2011 Oct;39(5):1185-90. doi: 10.1042/BST0391185. Biochem Soc Trans. 2011. PMID: 21936786 Review.
-
Clathrin-mediated endocytosis in budding yeast at a glance.J Cell Sci. 2016 Apr 15;129(8):1531-6. doi: 10.1242/jcs.182303. J Cell Sci. 2016. PMID: 27084361 Free PMC article. Review.
Cited by
-
Art2 mediates selective endocytosis of methionine transporters during adaptation to sphingolipid depletion.J Cell Sci. 2023 Jul 15;136(14):jcs260675. doi: 10.1242/jcs.260675. Epub 2023 Jul 25. J Cell Sci. 2023. PMID: 37337792 Free PMC article.
-
Spatial proteomics of vesicular trafficking: coupling mass spectrometry and imaging approaches in membrane biology.Plant Biotechnol J. 2023 Feb;21(2):250-269. doi: 10.1111/pbi.13929. Epub 2022 Nov 4. Plant Biotechnol J. 2023. PMID: 36204821 Free PMC article. Review.
-
A two-tiered system for selective receptor and transporter protein degradation.PLoS Genet. 2022 Oct 10;18(10):e1010446. doi: 10.1371/journal.pgen.1010446. eCollection 2022 Oct. PLoS Genet. 2022. PMID: 36215320 Free PMC article.
-
Methionine-triggered growth arrest reveals activation of Gcn2 by methionine transporter endocytosis.bioRxiv [Preprint]. 2025 May 14:2025.05.12.653625. doi: 10.1101/2025.05.12.653625. bioRxiv. 2025. PMID: 40462950 Free PMC article. Preprint.
References
-
- Ayscough K. R., Stryker J., Pokala N., Sanders M., Crews P., et al. , 1997. High rates of actin filament turnover in budding yeast and roles for actin in establishment and maintenance of cell polarity revealed using the actin inhibitor latrunculin-A. J. Cell Biol. 137(2): 399–416. 10.1083/jcb.137.2.399 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous