

Leukocyte integrin Mac-1 (CD11b/CD18, αMβ2, CR3) acts as a functional receptor for platelet factor 4
- PMID: 29540475
- PMCID: PMC5936813
- DOI: 10.1074/jbc.RA117.000515
Leukocyte integrin Mac-1 (CD11b/CD18, αMβ2, CR3) acts as a functional receptor for platelet factor 4
Abstract
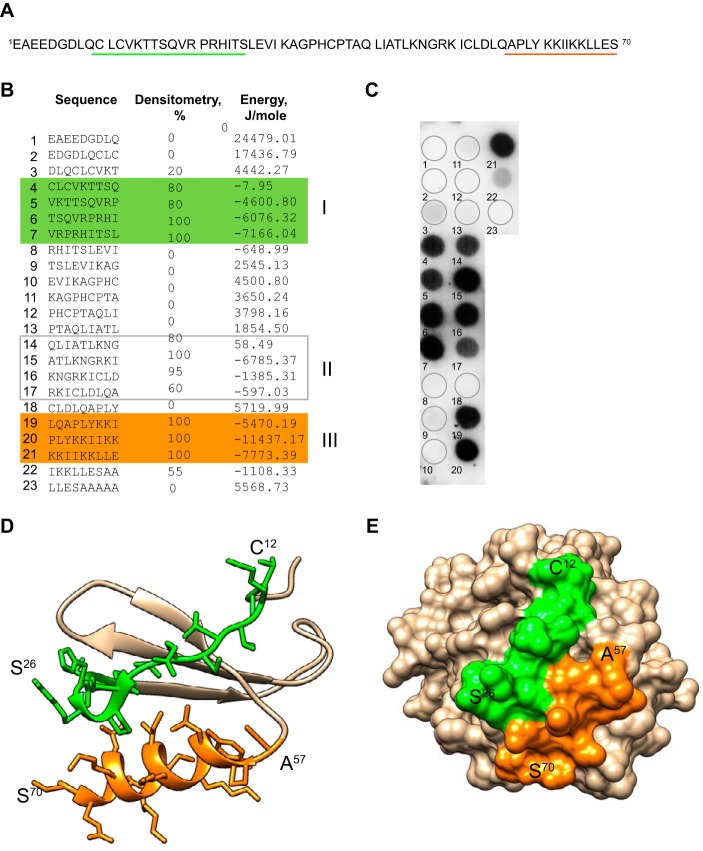
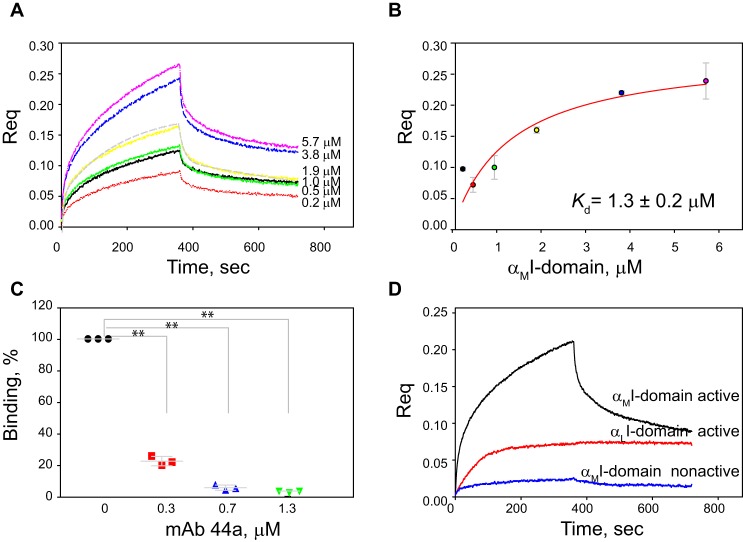
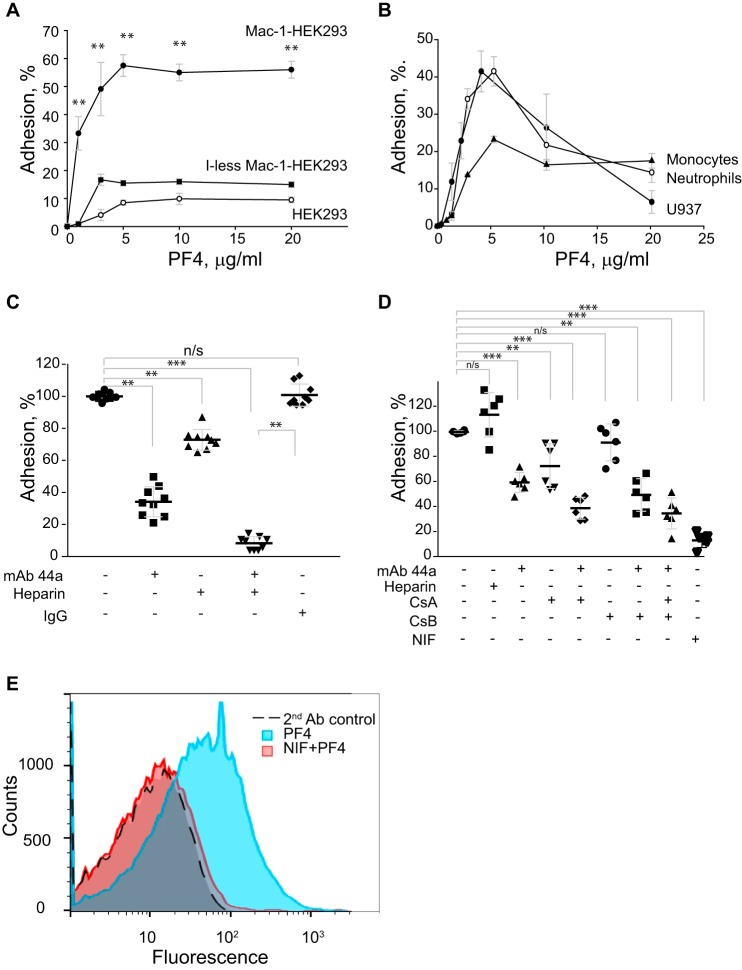
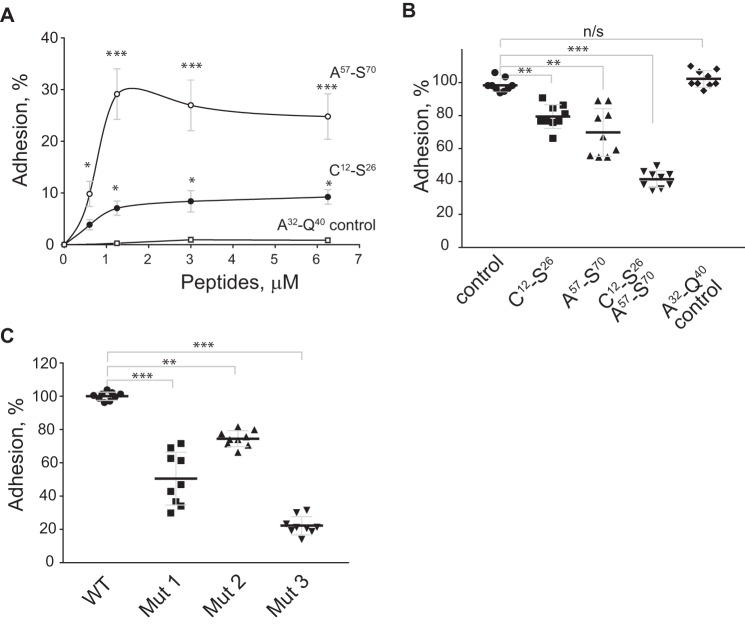
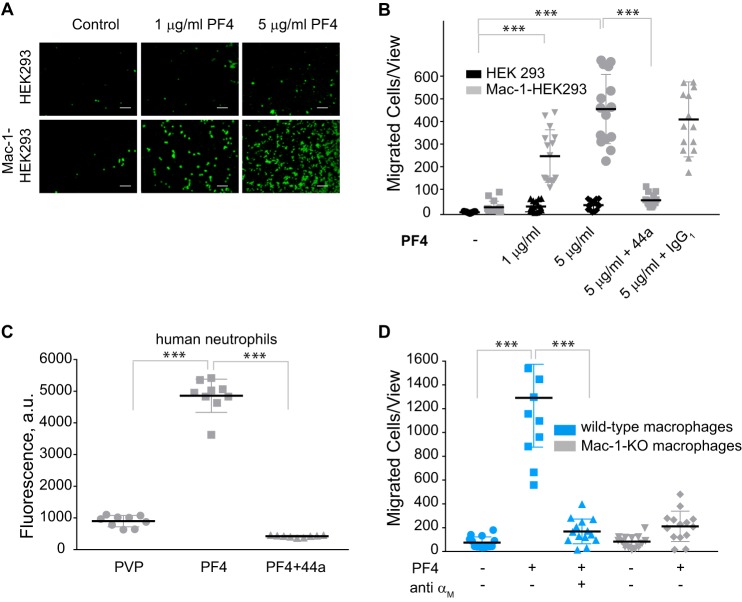
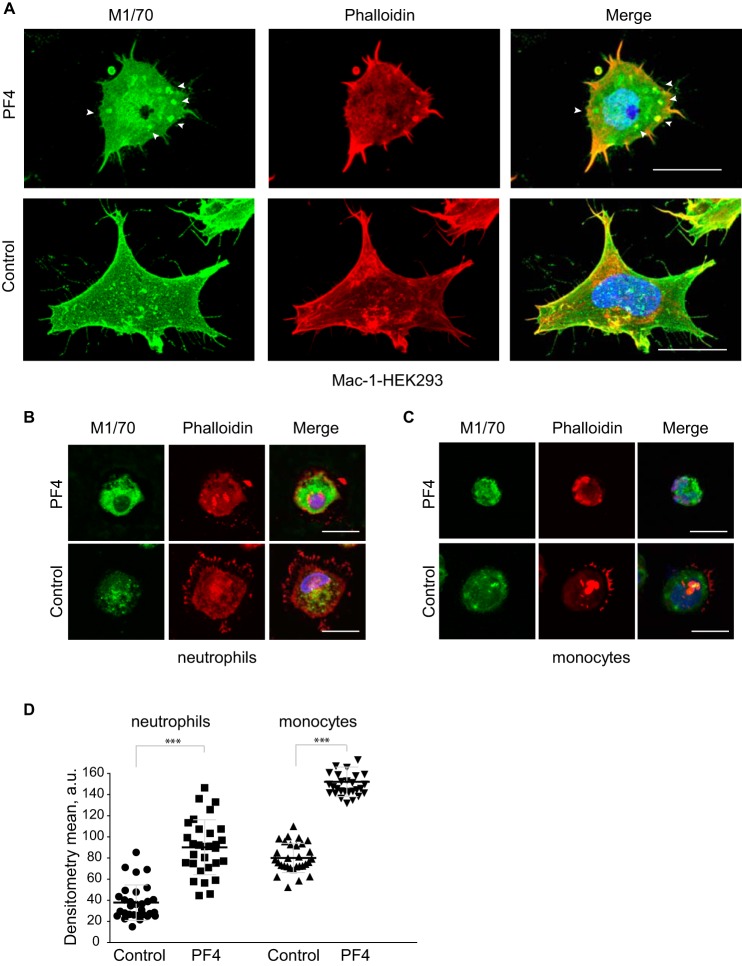
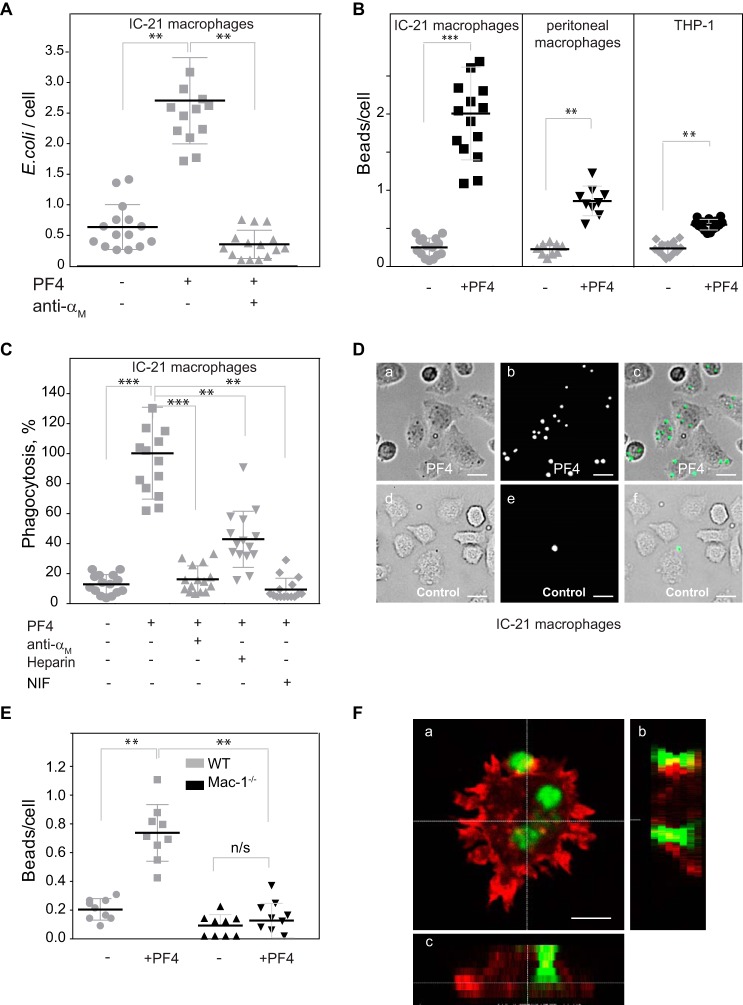
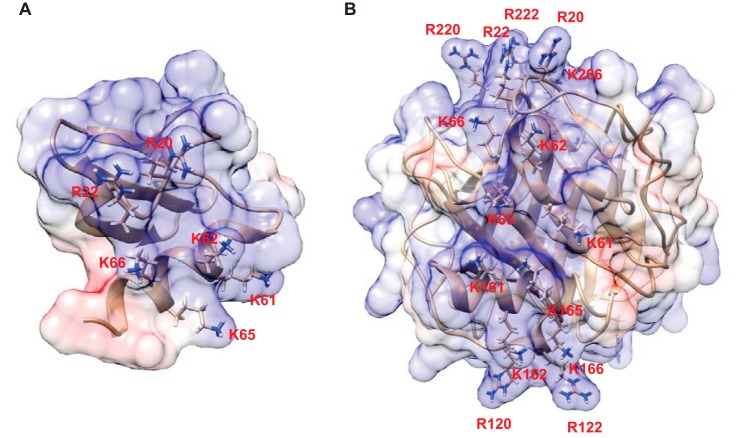
Platelet factor 4 (PF4) is one of the most abundant cationic proteins secreted from α-granules of activated platelets. Based on its structure, PF4 was assigned to the CXC family of chemokines and has been shown to have numerous effects on myeloid leukocytes. However, the receptor for PF4 remains unknown. Here, we demonstrate that PF4 induces leukocyte responses through the integrin Mac-1 (αMβ2, CD11b/CD18). Human neutrophils, monocytes, U937 monocytic and HEK293 cells expressing Mac-1 strongly adhered to immobilized PF4 in a concentration-dependent manner. The cell adhesion was partially blocked by anti-Mac-1 mAb and inhibition was enhanced when anti-Mac-1 antibodies were combined with glycosaminoglycans, suggesting that cell-surface proteoglycans act cooperatively with Mac-1. PF4 also induced Mac-1-dependent migration of human neutrophils and murine WT, but not Mac-1-deficient macrophages. Coating of Escherichia coli bacteria or latex beads with PF4 enhanced their phagocytosis by macrophages by ∼4-fold, and this process was blocked by different Mac-1 antagonists. Furthermore, PF4 potentiated phagocytosis by WT, but not Mac-1-deficient macrophages. As determined by biolayer interferometry, PF4 directly bound the αMI-domain, the major ligand-binding region of Mac-1, and this interaction was governed by a Kd of 1.3 ± 0.2 μm Using the PF4-derived peptide library, synthetic peptides duplicating the αMI-domain recognition sequences and recombinant mutant PF4 fragments, the binding sites for αMI-domain were identified in the PF4 segments Cys12-Ser26 and Ala57-Ser70 These results identify PF4 as a ligand for the integrin Mac-1 and suggest that many immune-modulating effects previously ascribed to PF4 are mediated through its interaction with Mac-1.
Keywords: Mac-1; PF4; alarmins; chemokine; integrin; macrophage; phagocytosis; platelet.
© 2018 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
Similar articles
-
Pleiotrophin, a multifunctional cytokine and growth factor, induces leukocyte responses through the integrin Mac-1.J Biol Chem. 2017 Nov 17;292(46):18848-18861. doi: 10.1074/jbc.M116.773713. Epub 2017 Sep 22. J Biol Chem. 2017. PMID: 28939773 Free PMC article.
-
The opioid peptide dynorphin A induces leukocyte responses via integrin Mac-1 (αMβ2, CD11b/CD18).Mol Pain. 2015 Jun 3;11:33. doi: 10.1186/s12990-015-0027-0. Mol Pain. 2015. PMID: 26036990 Free PMC article.
-
Identification of Human Cathelicidin Peptide LL-37 as a Ligand for Macrophage Integrin αMβ2 (Mac-1, CD11b/CD18) that Promotes Phagocytosis by Opsonizing Bacteria.Res Rep Biochem. 2016 Jul 7;2016(6):39-55. doi: 10.2147/rrbc.s107070. Res Rep Biochem. 2016. PMID: 27990411 Free PMC article.
-
Role of the lectin domain of Mac-1/CR3 (CD11b/CD18) in regulating intercellular adhesion.Immunol Res. 2002;25(3):219-27. doi: 10.1385/IR:25:3:219. Immunol Res. 2002. PMID: 12018461 Review.
-
The Promiscuous Profile of Complement Receptor 3 in Ligand Binding, Immune Modulation, and Pathophysiology.Front Immunol. 2021 Apr 29;12:662164. doi: 10.3389/fimmu.2021.662164. eCollection 2021. Front Immunol. 2021. PMID: 33995387 Free PMC article. Review.
Cited by
-
Staphylococcus aureus and Neutrophil Extracellular Traps: The Master Manipulator Meets Its Match in Immunothrombosis.Arterioscler Thromb Vasc Biol. 2022 Mar;42(3):261-276. doi: 10.1161/ATVBAHA.121.316930. Epub 2022 Feb 3. Arterioscler Thromb Vasc Biol. 2022. PMID: 35109674 Free PMC article. Review.
-
Platelet-neutrophil interaction in COVID-19 and vaccine-induced thrombotic thrombocytopenia.Front Immunol. 2023 May 19;14:1186000. doi: 10.3389/fimmu.2023.1186000. eCollection 2023. Front Immunol. 2023. PMID: 37275917 Free PMC article. Review.
-
αMI-domain of integrin Mac-1 binds the cytokine pleiotrophin using multiple mechanisms.Structure. 2024 Aug 8;32(8):1184-1196.e4. doi: 10.1016/j.str.2024.04.013. Epub 2024 May 9. Structure. 2024. PMID: 38729161 Free PMC article.
-
β2 Integrin Regulation of Neutrophil Functional Plasticity and Fate in the Resolution of Inflammation.Front Immunol. 2021 Mar 30;12:660760. doi: 10.3389/fimmu.2021.660760. eCollection 2021. Front Immunol. 2021. PMID: 33859651 Free PMC article. Review.
-
Role of platelet factor 4 in arteriovenous fistula maturation failure: What do we know so far?J Vasc Access. 2024 Mar;25(2):390-406. doi: 10.1177/11297298221085458. Epub 2022 Jun 24. J Vasc Access. 2024. PMID: 35751379 Free PMC article. Review.
References
-
- Afshar-Kharghan V., and Thiagarajan P. (2006) Leukocyte adhesion and thrombosis. Curr. Opin. Hematol. 13, 34–39 10.1097/01.moh.0000190107.54790.de - DOI - PubMed
-
- Hagberg I. A., Roald H. E., and Lyberg T. (1998) Adhesion of leukocytes to growing arterial thrombi. Thromb. Haemost. 80, 852–858 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
