

Bidirectional coordination of actions and habits by TrkB in mice
- PMID: 29540698
- PMCID: PMC5852142
- DOI: 10.1038/s41598-018-22560-x
Bidirectional coordination of actions and habits by TrkB in mice
Abstract
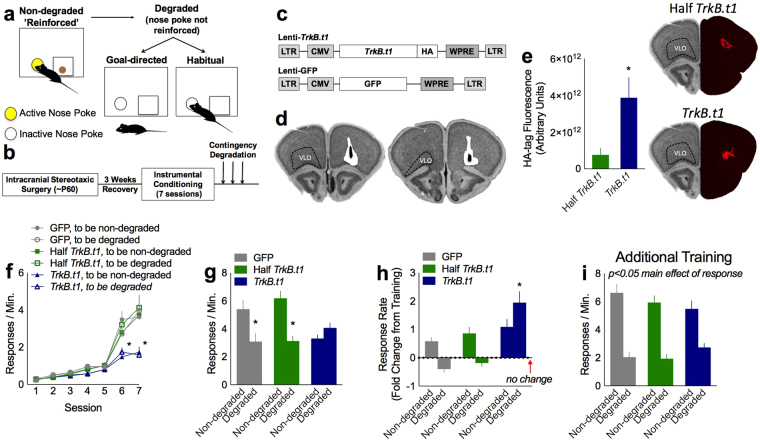
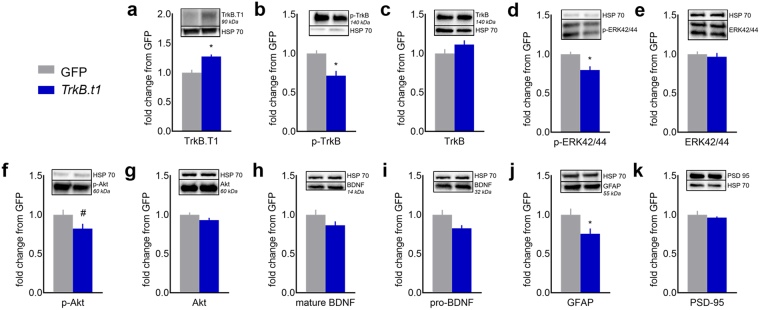
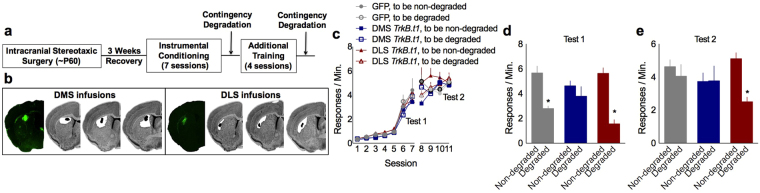
Specific corticostriatal structures and circuits are important for flexibly shifting between goal-oriented versus habitual behaviors. For example, the orbitofrontal cortex and dorsomedial striatum are critical for goal-directed action, while the dorsolateral striatum supports habits. To determine the role of neurotrophin signaling, we overexpressed a truncated, inactive form of tropomyosin receptor kinase B [also called tyrosine receptor kinase B (TrkB)], the high-affinity receptor for Brain-derived Neurotrophic Factor, in the orbitofrontal cortex, dorsomedial striatum and dorsolateral striatum. Overexpression of truncated TrkB interfered with phosphorylation of full-length TrkB and ERK42/44, as expected. In the orbitofrontal cortex and dorsomedial striatum, truncated trkB overexpression also occluded the ability of mice to select actions based on the likelihood that they would be reinforced. Meanwhile, in the dorsolateral striatum, truncated trkB blocked the development of habits. Thus, corticostriatal TrkB-mediated plasticity appears necessary for balancing actions and habits.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Action-Outcome Expectancies Require Orbitofrontal Neurotrophin Systems in Naïve and Cocaine-Exposed Mice.Neurotherapeutics. 2020 Jan;17(1):165-177. doi: 10.1007/s13311-019-00752-1. Neurotherapeutics. 2020. PMID: 31218603 Free PMC article.
-
NT-4/5 antagonizes the BDNF modulation of corticostriatal transmission: Role of the TrkB.T1 receptor.CNS Neurosci Ther. 2019 May;25(5):621-631. doi: 10.1111/cns.13091. Epub 2019 Jan 21. CNS Neurosci Ther. 2019. PMID: 30666798 Free PMC article.
-
Regulation of actions and habits by ventral hippocampal trkB and adolescent corticosteroid exposure.PLoS Biol. 2017 Nov 29;15(11):e2003000. doi: 10.1371/journal.pbio.2003000. eCollection 2017 Nov. PLoS Biol. 2017. PMID: 29186135 Free PMC article.
-
TrkB neurotrophin receptor at the core of antidepressant effects, but how?Cell Tissue Res. 2019 Jul;377(1):115-124. doi: 10.1007/s00441-018-02985-6. Epub 2019 Jan 12. Cell Tissue Res. 2019. PMID: 30637517 Review.
-
The significance of tyrosine kinase receptor B and brain-derived neurotrophic factor expression in salivary duct carcinoma.Ann Diagn Pathol. 2021 Feb;50:151673. doi: 10.1016/j.anndiagpath.2020.151673. Epub 2020 Nov 23. Ann Diagn Pathol. 2021. PMID: 33248386 Review.
Cited by
-
Identification of Novel BDNF-Specific Corticostriatal Circuitries.eNeuro. 2023 May 16;10(5):ENEURO.0238-21.2023. doi: 10.1523/ENEURO.0238-21.2023. Print 2023 May. eNeuro. 2023. PMID: 37156610 Free PMC article.
-
Control of goal-directed and inflexible actions by dorsal striatal melanocortin systems, in coordination with the central nucleus of the amygdala.Prog Neurobiol. 2024 Jul;238:102629. doi: 10.1016/j.pneurobio.2024.102629. Epub 2024 May 17. Prog Neurobiol. 2024. PMID: 38763506 Free PMC article.
-
Cocaine disrupts action flexibility via glucocorticoid receptors.iScience. 2024 May 28;27(7):110148. doi: 10.1016/j.isci.2024.110148. eCollection 2024 Jul 19. iScience. 2024. PMID: 38989467 Free PMC article.
-
A Truncated Receptor TrkB Isoform (TrkB.T1) in Mechanisms of Genetically Determined Depressive-like Behavior of Mice.Biomedicines. 2023 Sep 19;11(9):2573. doi: 10.3390/biomedicines11092573. Biomedicines. 2023. PMID: 37761014 Free PMC article.
-
Persistence of learning-induced synapses depends on neurotrophic priming of glucocorticoid receptors.Proc Natl Acad Sci U S A. 2019 Jun 25;116(26):13097-13106. doi: 10.1073/pnas.1903203116. Epub 2019 Jun 10. Proc Natl Acad Sci U S A. 2019. PMID: 31182610 Free PMC article.
References
-
- Balleine BW, O’Doherty JP. Human and rodent homologies in action control: corticostriatal determinants of goal-directed and habitual action. Neuropsychopharmacology: official publication of the American College of Neuropsychopharmacology. 2010;35:48–69. doi: 10.1038/npp.2009.131. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
