

Voltage gating of mechanosensitive PIEZO channels
- PMID: 29545531
- PMCID: PMC5854696
- DOI: 10.1038/s41467-018-03502-7
Voltage gating of mechanosensitive PIEZO channels
Abstract
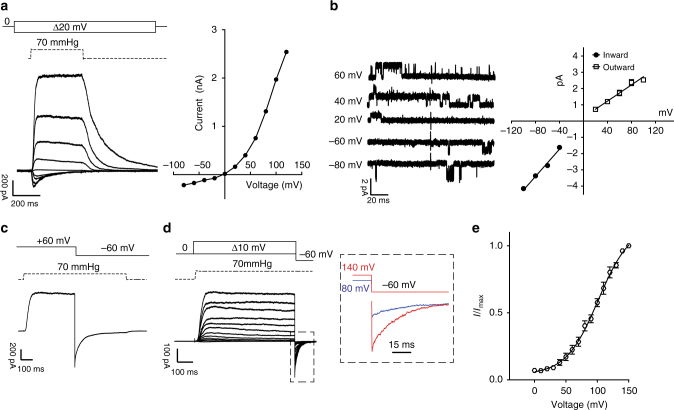
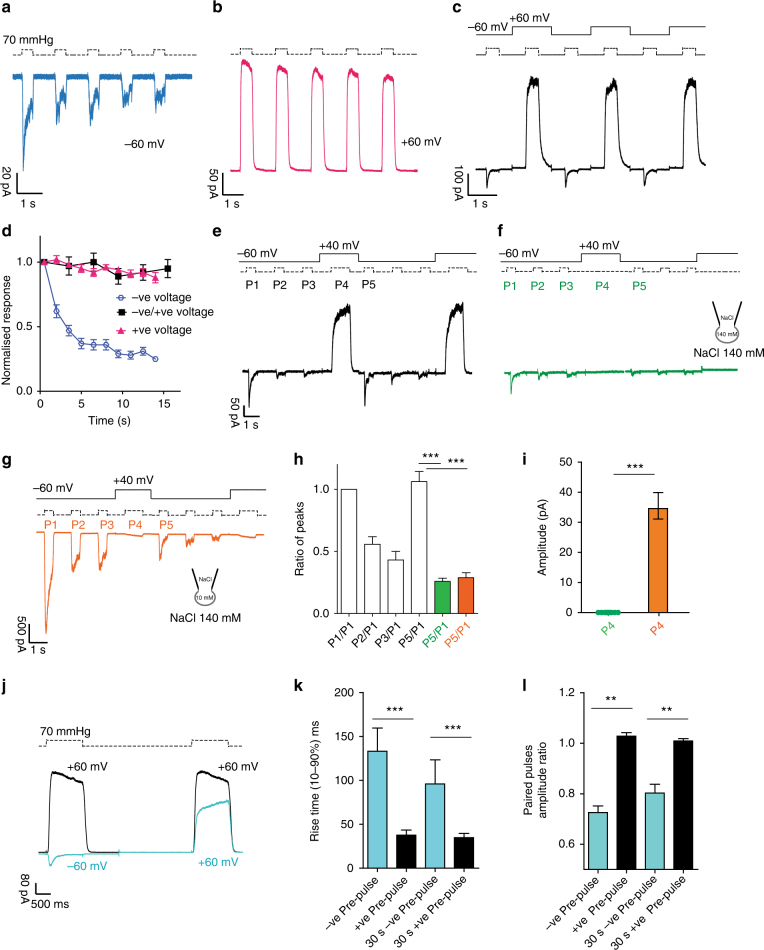
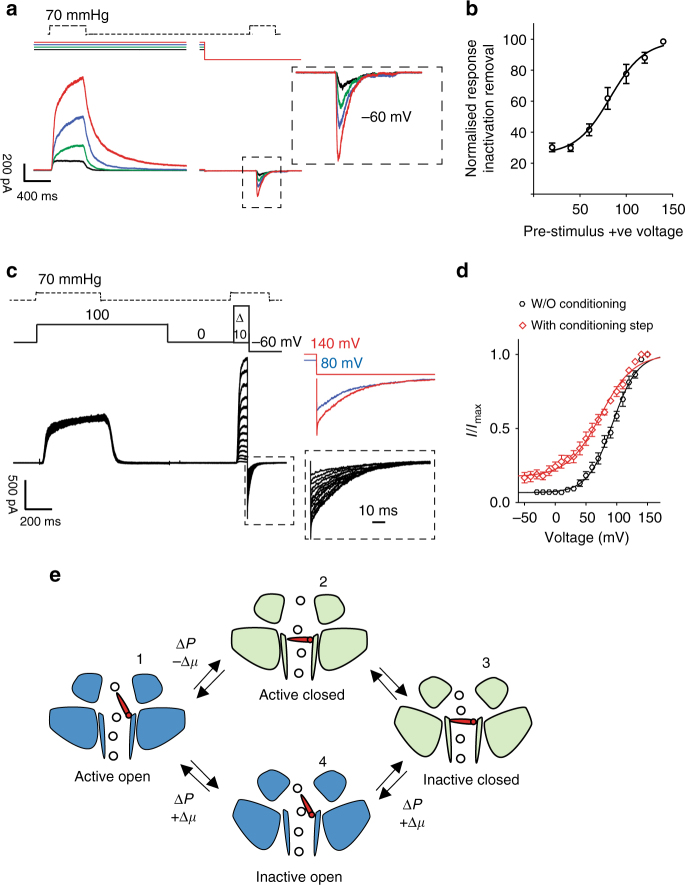
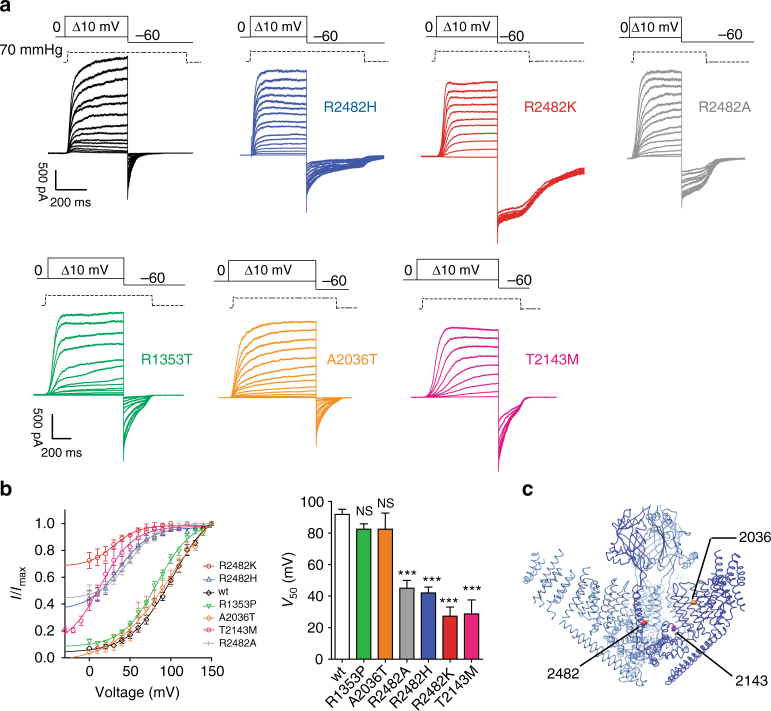
Mechanosensitive PIEZO ion channels are evolutionarily conserved proteins whose presence is critical for normal physiology in multicellular organisms. Here we show that, in addition to mechanical stimuli, PIEZO channels are also powerfully modulated by voltage and can even switch to a purely voltage-gated mode. Mutations that cause human diseases, such as xerocytosis, profoundly shift voltage sensitivity of PIEZO1 channels toward the resting membrane potential and strongly promote voltage gating. Voltage modulation may be explained by the presence of an inactivation gate in the pore, the opening of which is promoted by outward permeation. Older invertebrate (fly) and vertebrate (fish) PIEZO proteins are also voltage sensitive, but voltage gating is a much more prominent feature of these older channels. We propose that the voltage sensitivity of PIEZO channels is a deep property co-opted to add a regulatory mechanism for PIEZO activation in widely different cellular contexts.
Conflict of interest statement
The authors declare no competing interests.
Figures
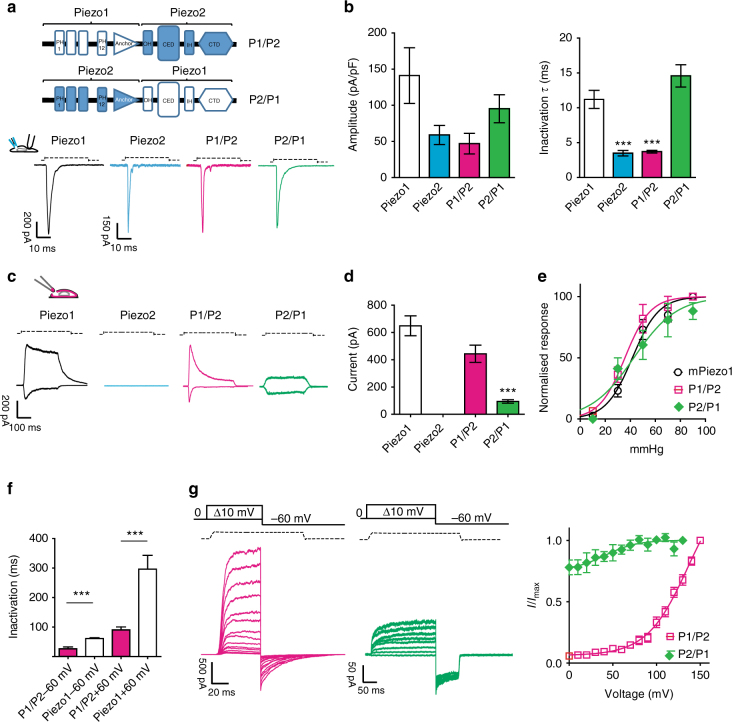
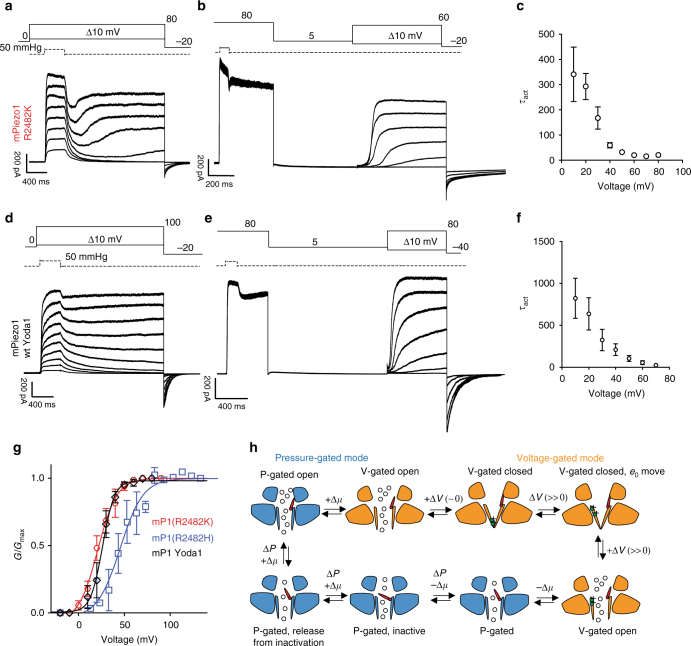
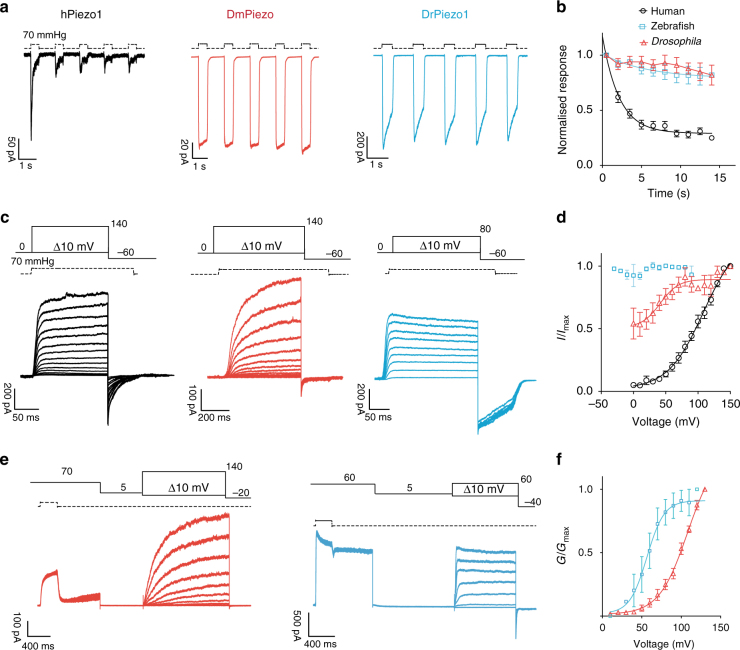
Comment in
-
Commentary: Voltage Gating of Mechanosensitive PIEZO Channels.Front Physiol. 2018 Nov 20;9:1565. doi: 10.3389/fphys.2018.01565. eCollection 2018. Front Physiol. 2018. PMID: 30524293 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
