

Solution NMR of SNAREs, complexin and α-synuclein in association with membrane-mimetics
- PMID: 29548366
- PMCID: PMC5863748
- DOI: 10.1016/j.pnmrs.2018.02.001
Solution NMR of SNAREs, complexin and α-synuclein in association with membrane-mimetics
Abstract
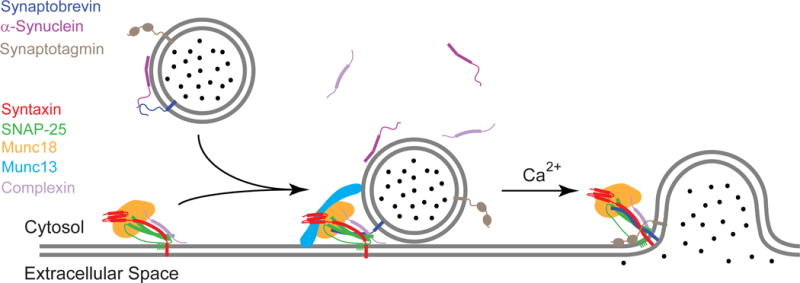
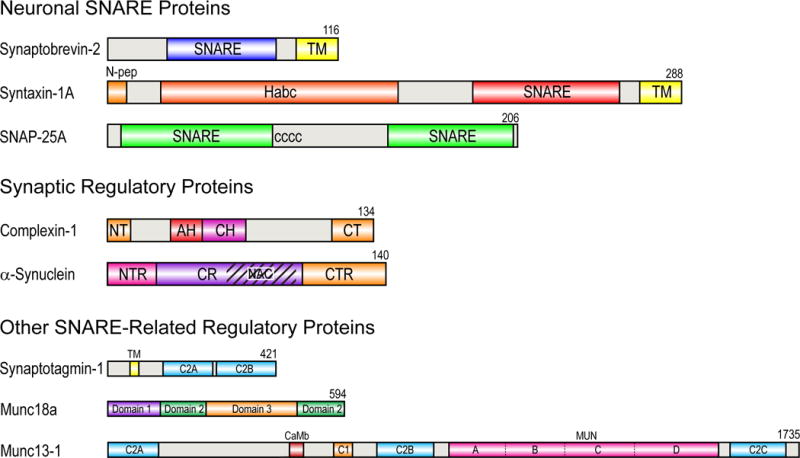
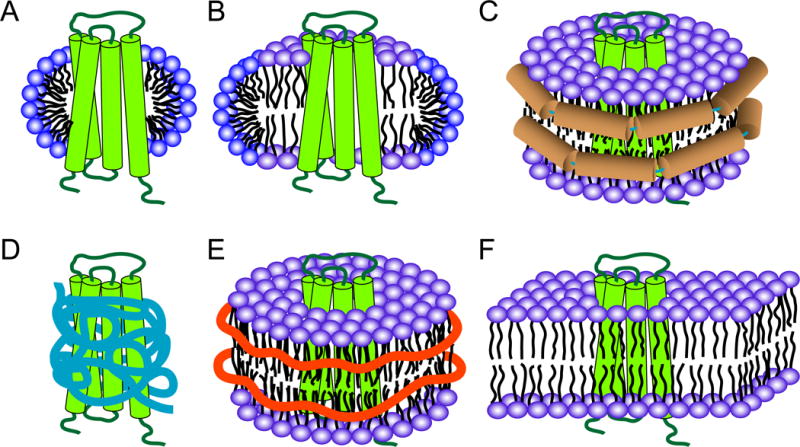
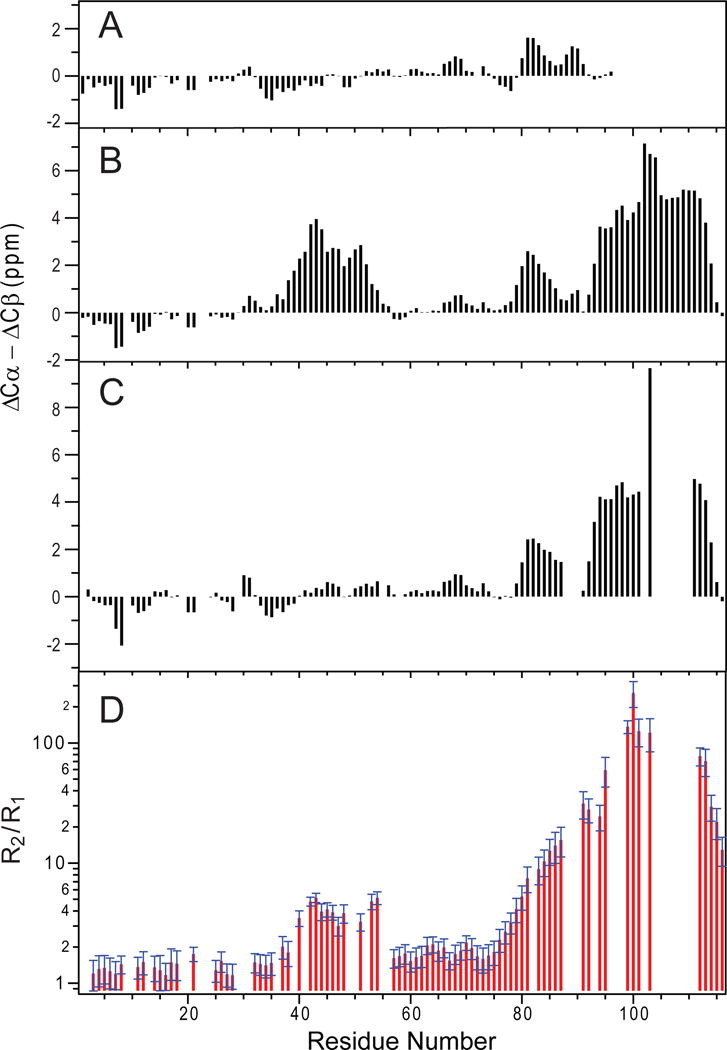
SNARE-mediated membrane fusion is a ubiquitous process responsible for intracellular vesicle trafficking, including membrane fusion in exocytosis that leads to hormone and neurotransmitter release. The proteins that facilitate this process are highly dynamic and adopt multiple conformations when they interact with other proteins and lipids as they form highly regulated molecular machines that operate on membranes. Solution NMR is an ideal method to capture high-resolution glimpses of the molecular transformations that take place when these proteins come together and work on membranes. Since solution NMR has limitations on the size of proteins and complexes that can be studied, lipid bilayer model membranes cannot be used in these approaches, so the relevant interactions are typically studied in various types of membrane-mimetics that are tractable by solution NMR methods. In this review we therefore first summarize different membrane-mimetic systems that are commonly used or that show promise for solution NMR studies of membrane-interacting proteins. We then summarize recent NMR studies on two SNARE proteins, syntaxin and synaptobrevin, and two related regulatory proteins, complexin and α-synuclein, and their interactions with membrane lipids. These studies provide a structural and dynamical framework for how these proteins might carry out their functions in the vicinity of lipid membranes. The common theme throughout these studies is that membrane interactions have major influences on the structural dynamics of these proteins that cannot be ignored when attempting to explain their functions in contemporary models of SNARE-mediated membrane fusion.
Copyright © 2018 Elsevier B.V. All rights reserved.
Figures
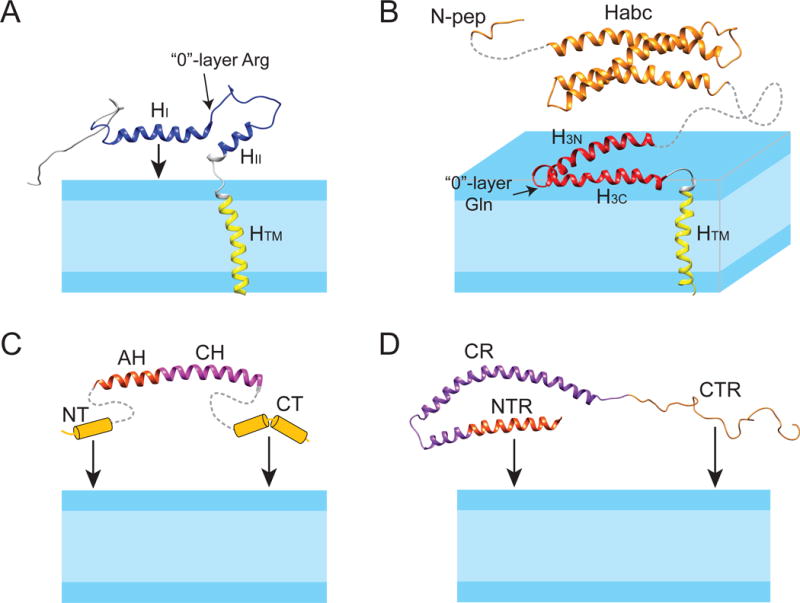
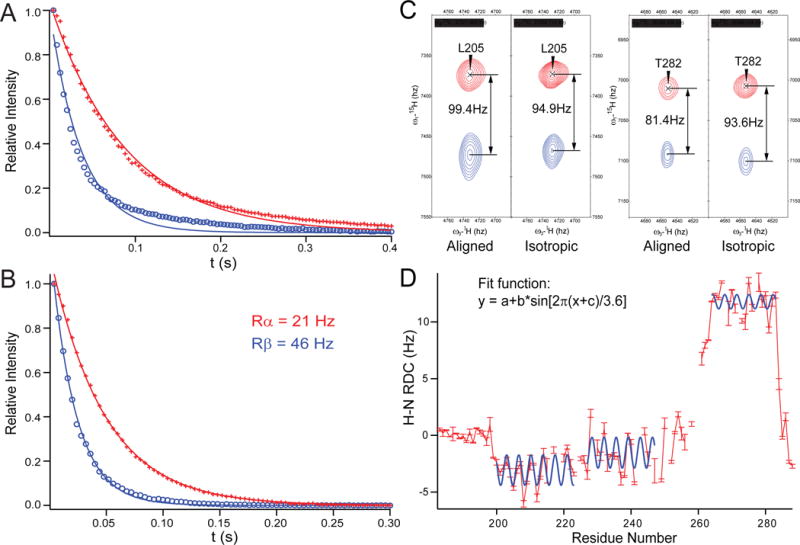
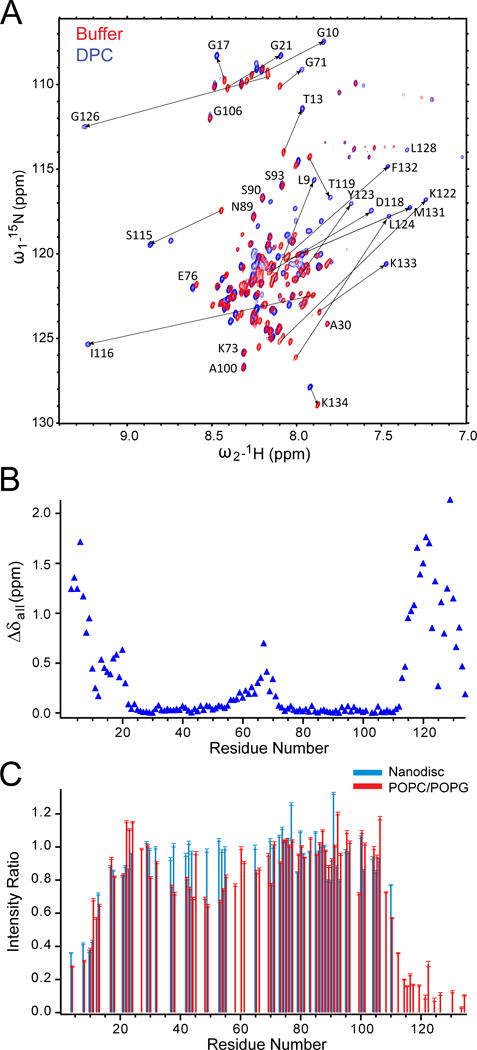
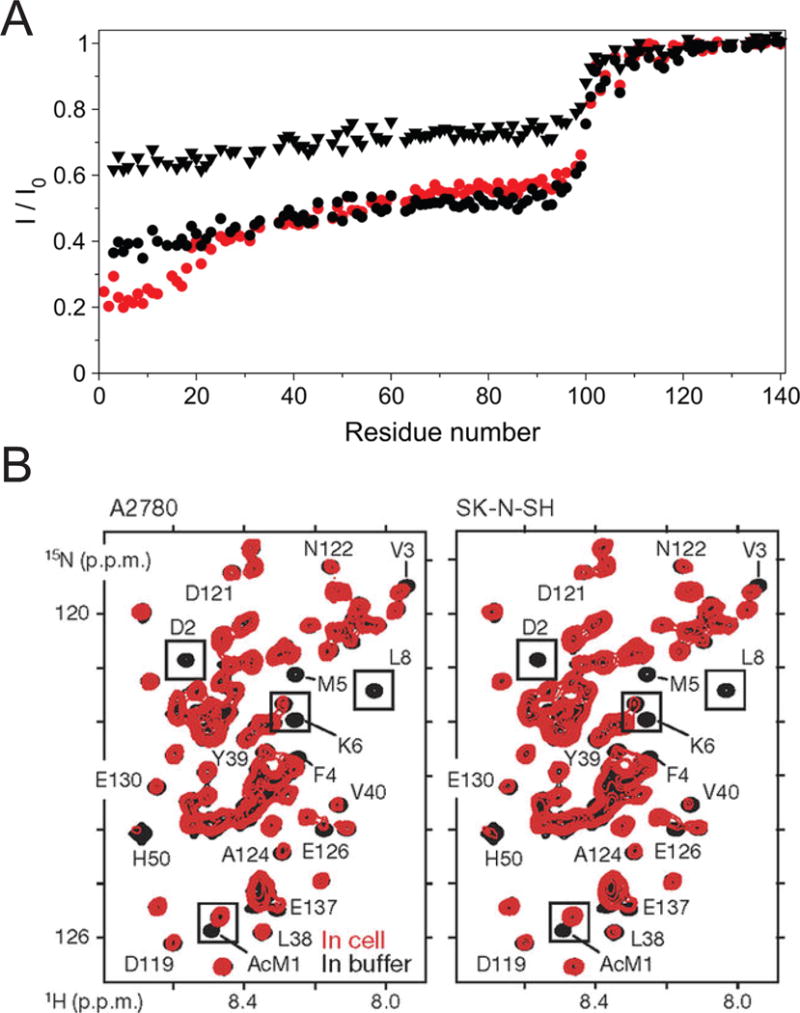
Similar articles
-
Reconciling isothermal titration calorimetry analyses of interactions between complexin and truncated SNARE complexes.Elife. 2017 Sep 7;6:e30286. doi: 10.7554/eLife.30286. Elife. 2017. PMID: 28880148 Free PMC article.
-
Interaction of the Complexin Accessory Helix with Synaptobrevin Regulates Spontaneous Fusion.Biophys J. 2016 Nov 1;111(9):1954-1964. doi: 10.1016/j.bpj.2016.09.017. Biophys J. 2016. PMID: 27806277 Free PMC article.
-
Complexin induces a conformational change at the membrane-proximal C-terminal end of the SNARE complex.Elife. 2016 Jun 2;5:e16886. doi: 10.7554/eLife.16886. Elife. 2016. PMID: 27253060 Free PMC article.
-
Molecular Mechanisms Underlying Neurotransmitter Release.Annu Rev Biophys. 2022 May 9;51:377-408. doi: 10.1146/annurev-biophys-111821-104732. Epub 2022 Feb 15. Annu Rev Biophys. 2022. PMID: 35167762 Free PMC article. Review.
-
For better or for worse: complexins regulate SNARE function and vesicle fusion.Traffic. 2008 Sep;9(9):1403-13. doi: 10.1111/j.1600-0854.2008.00758.x. Epub 2008 Apr 28. Traffic. 2008. PMID: 18445121 Review.
Cited by
-
The Influence of the Mixed DPC:SDS Micelle on the Structure and Oligomerization Process of the Human Cystatin C.Membranes (Basel). 2020 Dec 24;11(1):17. doi: 10.3390/membranes11010017. Membranes (Basel). 2020. PMID: 33374409 Free PMC article.
-
Visiting order on membrane proteins by using nanotechnology.Proc Natl Acad Sci U S A. 2007 Apr 17;104(16):6502-3. doi: 10.1073/pnas.0701950104. Epub 2007 Apr 9. Proc Natl Acad Sci U S A. 2007. PMID: 17420448 Free PMC article. No abstract available.
-
The effects of biological crowders on fibrillization, structure, diffusion, and conformational dynamics of α-synuclein.Protein Sci. 2024 Mar;33(3):e4894. doi: 10.1002/pro.4894. Protein Sci. 2024. PMID: 38358134 Free PMC article.
-
Detergent-mediated protein aggregation.Chem Phys Lipids. 2013 Apr;169:72-84. doi: 10.1016/j.chemphyslip.2013.02.005. Epub 2013 Mar 4. Chem Phys Lipids. 2013. PMID: 23466535 Free PMC article. Review.
-
Isotopic Labeling of Eukaryotic Membrane Proteins for NMR Studies of Interactions and Dynamics.Methods Enzymol. 2019;614:37-65. doi: 10.1016/bs.mie.2018.08.030. Epub 2018 Dec 18. Methods Enzymol. 2019. PMID: 30611431 Free PMC article.
References
-
- Weber T, Zemelman BV, McNew JA, Westermann B, Gmachl M, Parlati F, Sollner TH, Rothman JE. SNAREpins: Minimal machinery for membrane fusion. Cell. 1998;92:759–772. - PubMed
-
- Jahn R, Scheller RH. SNAREs–engines for membrane fusion. Nat Rev Mol Cell Biol. 2006;7:631–643. - PubMed
-
- Sutton RB, Fasshauer D, Jahn R, Brunger AT. Crystal structure of a SNARE complex involved in synaptic exocytosis at 2.4 A resolution. Nature. 1998;395:347–353. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
