

In vitro exploration of ACAT contributions to lipid droplet formation during adipogenesis
- PMID: 29549095
- PMCID: PMC5928425
- DOI: 10.1194/jlr.M081745
In vitro exploration of ACAT contributions to lipid droplet formation during adipogenesis
Abstract
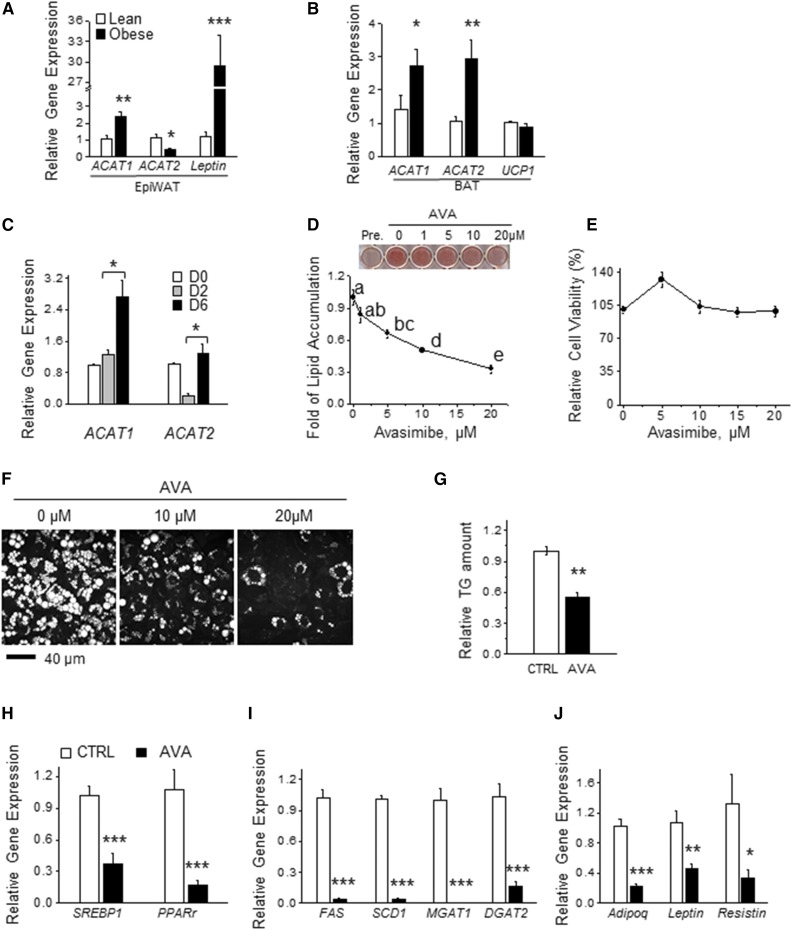
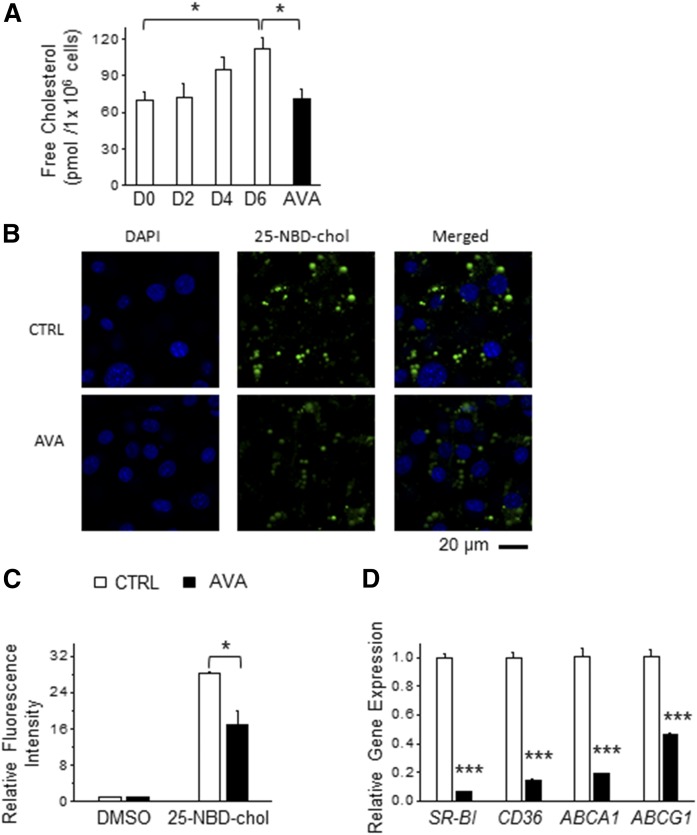
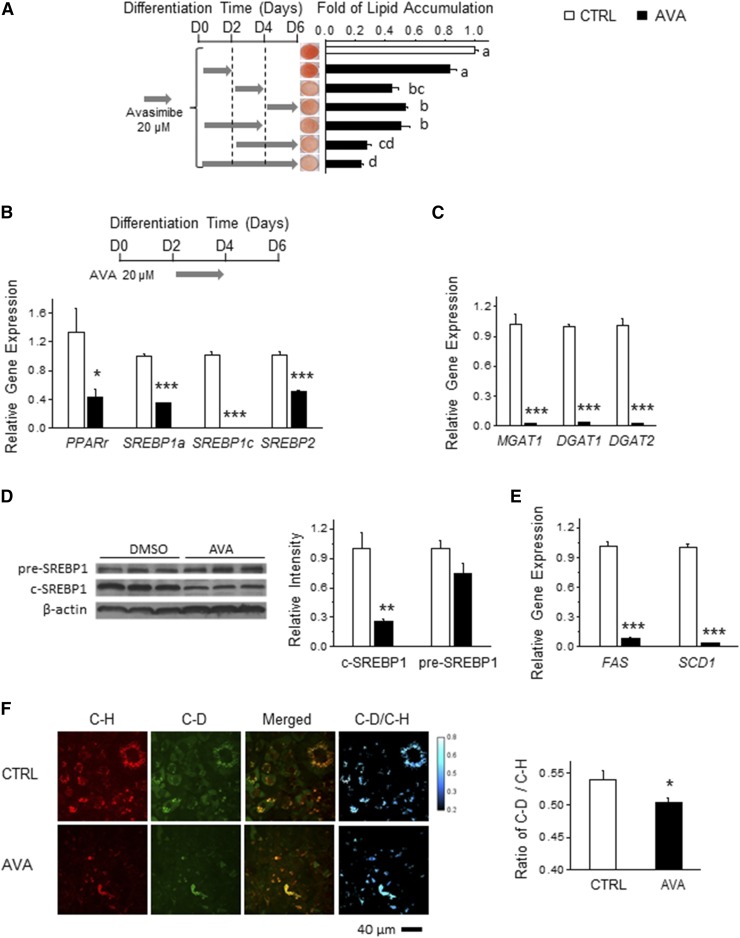
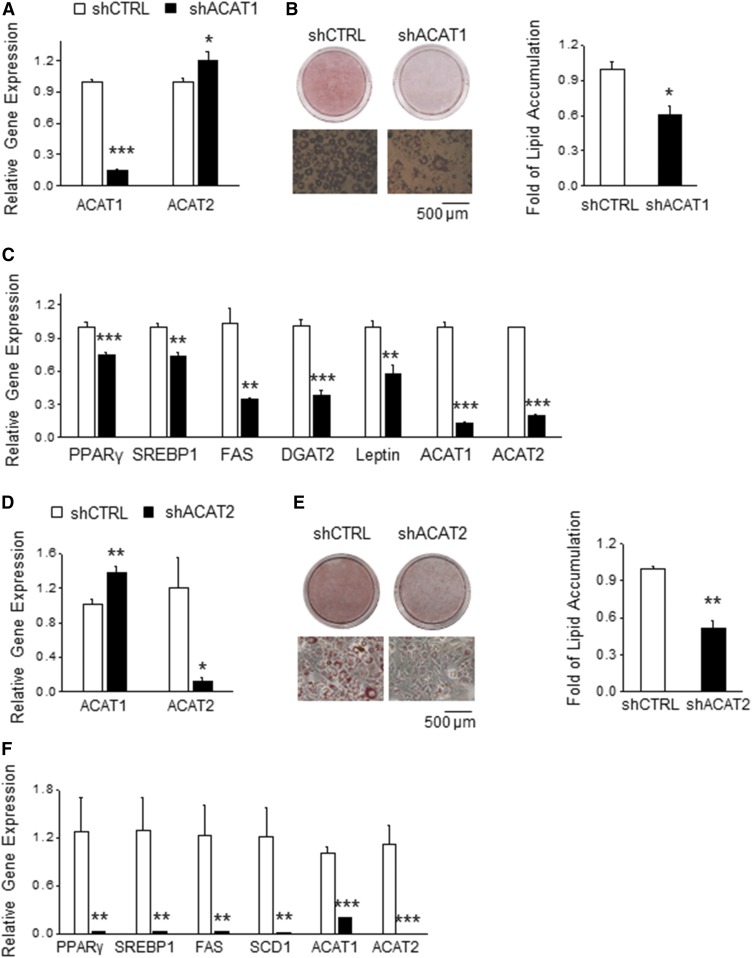
As adipose tissue is the major cholesterol storage organ and most of the intracellular cholesterol is distributed to lipid droplets (LDs), cholesterol homeostasis may have a role in the regulation of adipocyte size and function. ACATs catalyze the formation of cholesteryl ester (CE) from free cholesterol to modulate the cholesterol balance. Despite the well-documented role of ACATs in hypercholesterolemia, their role in LD development during adipogenesis remains elusive. Here, we identify ACATs as regulators of de novo lipogenesis and LD formation in murine 3T3-L1 adipocytes. Pharmacological inhibition of ACAT activity suppressed intracellular cholesterol and CE levels, and reduced expression of genes involved in cholesterol uptake and efflux. ACAT inhibition resulted in decreased de novo lipogenesis, as demonstrated by reduced maturation of SREBP1 and SREBP1-downstream lipogenic gene expression. Consistent with this observation, knockdown of either ACAT isoform reduced total adipocyte lipid content by approximately 40%. These results demonstrate that ACATs are required for storage ability of lipids and cholesterol in adipocytes.
Keywords: acyl-CoA:cholesterol acyltransferase; adipocytes; avasimibe; cholesterol metabolism; fatty acid synthesis; triglycerides.
Copyright © 2018 by the American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The other authors declare no conflicts of interest.
Figures
References
-
- Farkas J., Angel A., and Avigan M. I.. 1973. Studies on the compartmentation of lipid in adipose cells. II. Cholesterol accumulation and distribution in adipose tissue components. J. Lipid Res. 14: 344–356. - PubMed
-
- Prattes S., Horl G., Hammer A., Blaschitz A., Graier W. F., Sattler W., Zechner R., and Steyrer E.. 2000. Intracellular distribution and mobilization of unesterified cholesterol in adipocytes: triglyceride droplets are surrounded by cholesterol-rich ER-like surface layer structures. J. Cell Sci. 113: 2977–2989. - PubMed
-
- Le Lay S., Krief S., Farnier C., Lefrere I., Le Liepvre X., Bazin R., Ferre P., and Dugail I.. 2001. Cholesterol, a cell size-dependent signal that regulates glucose metabolism and gene expression in adipocytes. J. Biol. Chem. 276: 16904–16910. - PubMed
-
- Kovanen P. T., Nikkila E. A., and Miettinen T. A.. 1975. Regulation of cholesterol synthesis and storage in fat cells. J. Lipid Res. 16: 211–223. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
