

Genome-Wide Analysis of Corynespora cassiicola Leaf Fall Disease Putative Effectors
- PMID: 29551995
- PMCID: PMC5840194
- DOI: 10.3389/fmicb.2018.00276
Genome-Wide Analysis of Corynespora cassiicola Leaf Fall Disease Putative Effectors
Abstract
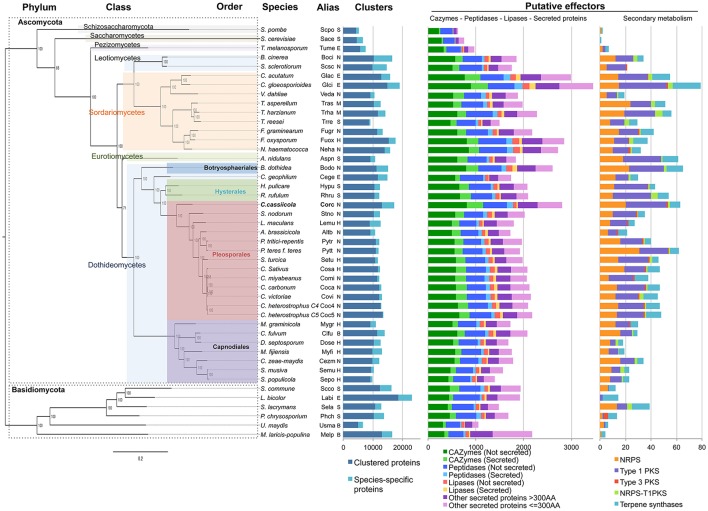
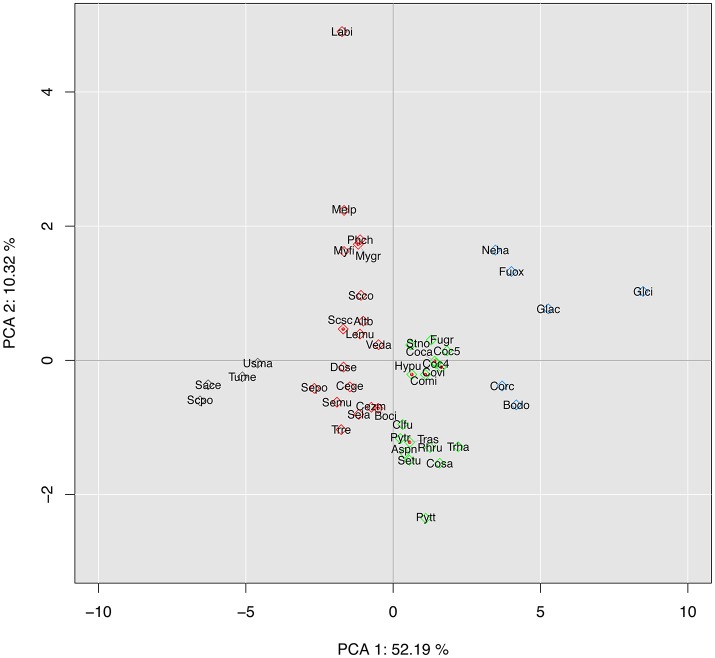
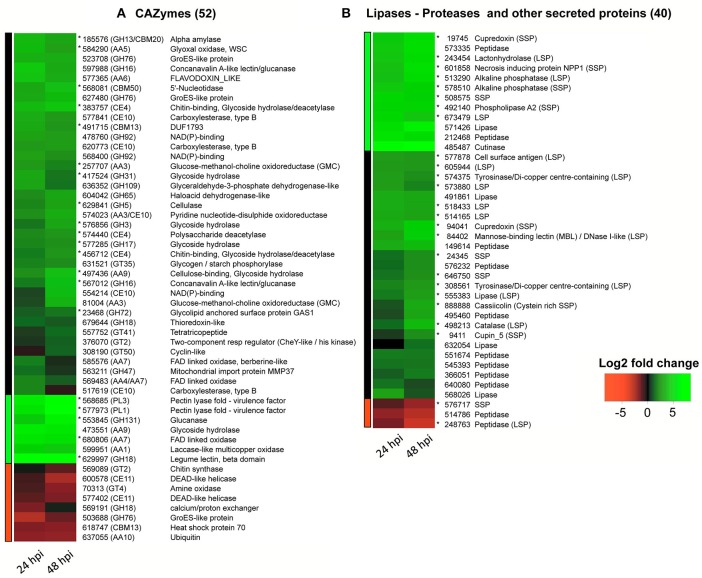
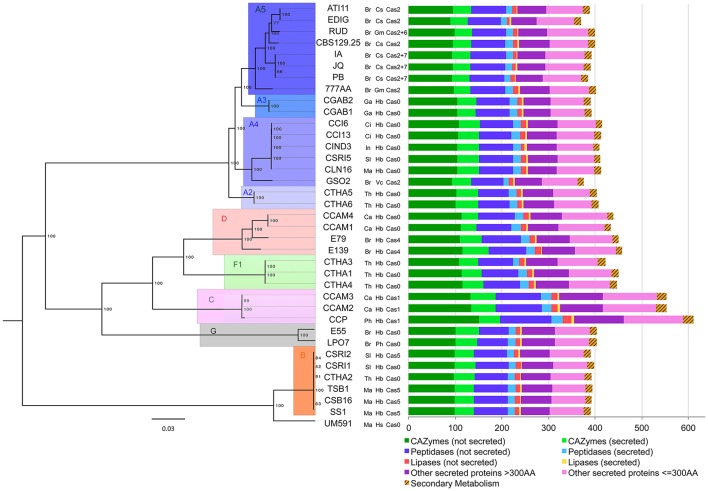
Corynespora cassiicola is an Ascomycetes fungus with a broad host range and diverse life styles. Mostly known as a necrotrophic plant pathogen, it has also been associated with rare cases of human infection. In the rubber tree, this fungus causes the Corynespora leaf fall (CLF) disease, which increasingly affects natural rubber production in Asia and Africa. It has also been found as an endophyte in South American rubber plantations where no CLF outbreak has yet occurred. The C. cassiicola species is genetically highly diverse, but no clear relationship has been evidenced between phylogenetic lineage and pathogenicity. Cassiicolin, a small glycosylated secreted protein effector, is thought to be involved in the necrotrophic interaction with the rubber tree but some virulent C. cassiicola isolates do not have a cassiicolin gene. This study set out to identify other putative effectors involved in CLF. The genome of a highly virulent C. cassiicola isolate from the rubber tree (CCP) was sequenced and assembled. In silico prediction revealed 2870 putative effectors, comprising CAZymes, lipases, peptidases, secreted proteins and enzymes associated with secondary metabolism. Comparison with the genomes of 44 other fungal species, focusing on effector content, revealed a striking proximity with phylogenetically unrelated species (Colletotrichum acutatum, Colletotrichum gloesporioides, Fusarium oxysporum, nectria hematococca, and Botrosphaeria dothidea) sharing life style plasticity and broad host range. Candidate effectors involved in the compatible interaction with the rubber tree were identified by transcriptomic analysis. Differentially expressed genes included 92 putative effectors, among which cassiicolin and two other secreted singleton proteins. Finally, the genomes of 35 C. cassiicola isolates representing the genetic diversity of the species were sequenced and assembled, and putative effectors identified. At the intraspecific level, effector-based classification was found to be highly consistent with the phylogenomic trees. Identification of lineage-specific effectors is a key step toward understanding C. cassiicola virulence and host specialization mechanisms.
Keywords: Corynespora cassiicola; Hevea brasiliensis; cassiicolin; effectors; gene expression; genomics; plant-pathogens interaction.
Figures

References
-
- Atan S., Hamid N. H. (2003). Differentiating races of Corynespora cassiicola using RAPD and internal transcribed spacer markers. J. Rubber Res. 6, 58–64.
-
- Benjamini Y., Yekutieli D. (2001). The control of the false discovery rate in multiple testing under dependency. Ann. Stat. 29, 1165–1188. 10.1214/aos/1013699998 - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous