

Mechanistic Studies of 1-Deoxy-D-Xylulose-5-Phosphate Synthase from Deinococcus radiodurans
- PMID: 29552677
- PMCID: PMC5851014
- DOI: 10.21767/2471-8084.100051
Mechanistic Studies of 1-Deoxy-D-Xylulose-5-Phosphate Synthase from Deinococcus radiodurans
Abstract
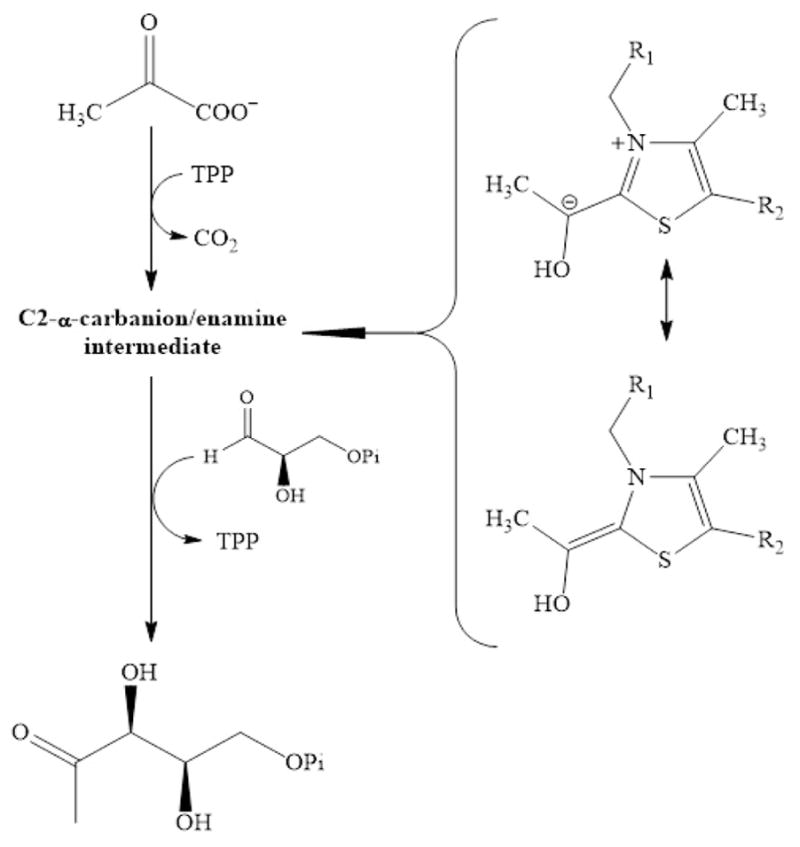
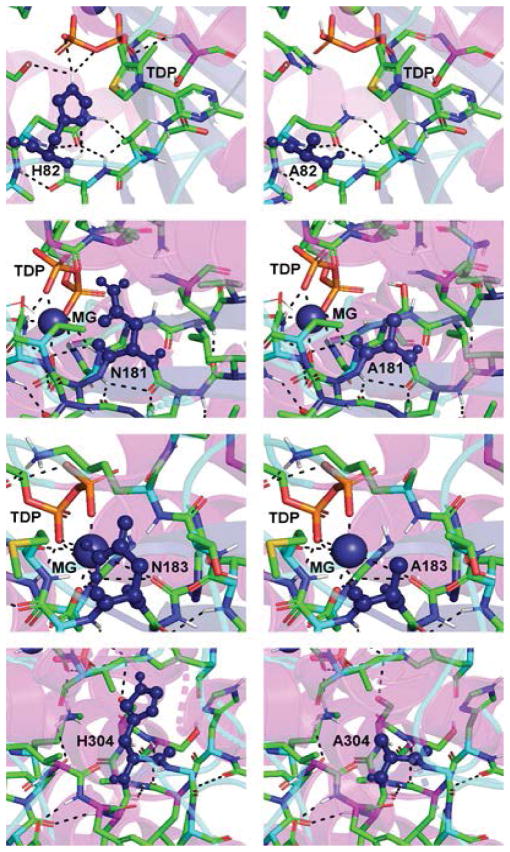
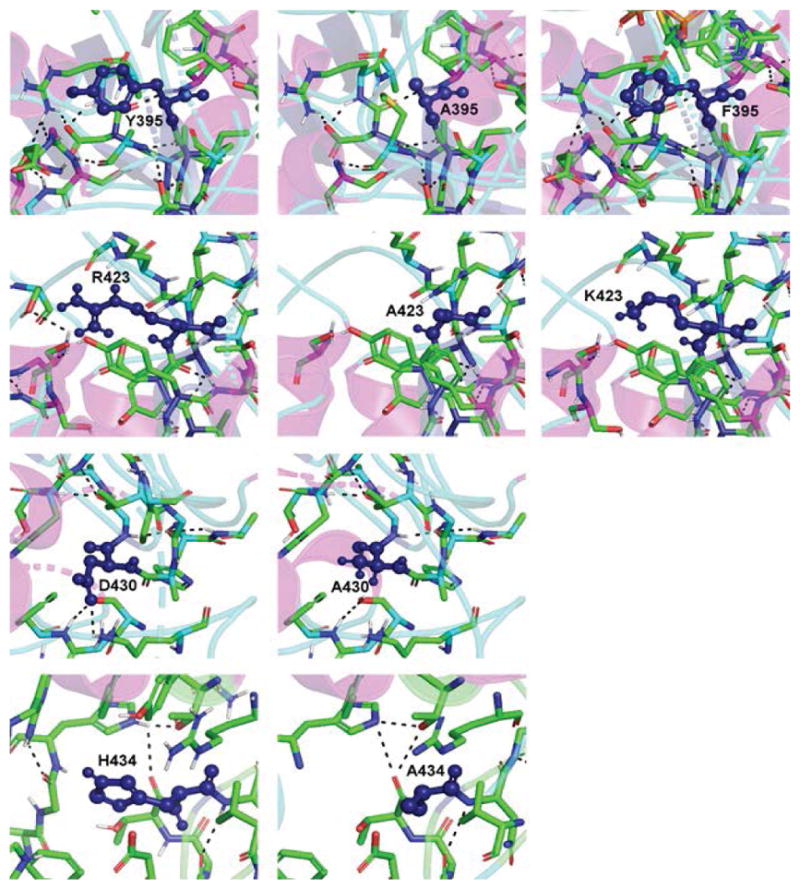
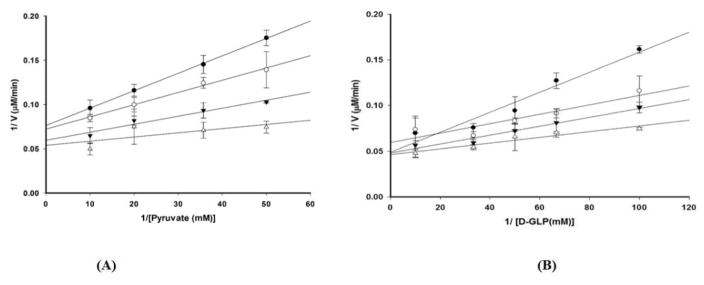
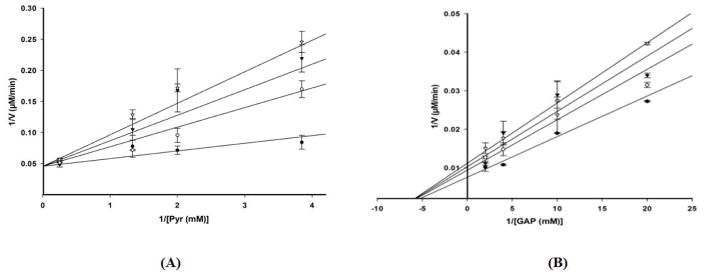
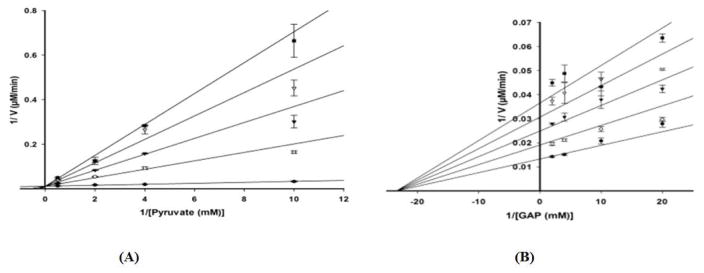
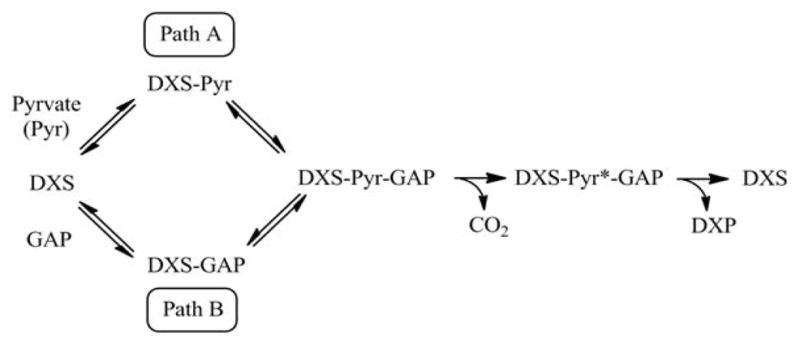
The non-mevalonate dependent (NMVA) pathway for the biosynthesis of isopentenyl pyrophosphate and dimethylallyl pyrophosphate is the sole source of these terpenoids for the production of isoprenoids in the apicomplexan parasites, in many eubacteria, and in plants. The absence of this pathway in higher organisms has opened a new platform for the development of novel antibiotics and anti-malarials. The enzyme catalyzing the first step of the NMVA pathway is 1-deoxy-D-xylulose-5-phosphate synthase (DXPS). DXPS catalyzes the thiamine pyrophosphate- and Mg (II)-dependent conjugation of pyruvate and D-glyceraldehyde-3-phosphate to form 1-deoxy-D-xylulose-5-phosphate and CO2. The kinetic mechanism of DXPS from Deinococcus radiodurans most consistent with our data is random sequential as shown using a combination of kinetic analysis and product and dead-end inhibition studies. The role of active site amino acids, identified by sequence alignment to other DXPS proteins, was probed by constructing and analyzing the catalytic efficacy of a set of targeted site-directed mutants.
Keywords: Dimethylallyl pyrophosphate; Isopentenyl pyrophosphate; Non-mevalonate; Site-directed mutagenesis; TPP-dependent; α-Carbanion/Enamine intermediate.
Figures
References
-
- Friedman M, Byers SO, St George S. Cholesterol metabolism. Ann Rev Biochem. 1956;25:613–640. - PubMed
-
- Popják G. Biosynthesis of cholesterol and related substances. Annu Rev Biochem. 1958;27:533–560. - PubMed
-
- Cornforth JW. Biosynthesis of fatty acids and cholesterol considered as chemical processes. J Lipid Res. 1959;1:3–28.
-
- Bucher NLR, Overath P, Lynen F. Beta-hydroxy-beta-methylglutaryl coenzyme A reductase, cleavage and condensing enzymes in relation to cholesterol formation in rat liver. Biochim Biophys Acta. 1960;40:491–501. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources