

A human monoclonal antibody prevents malaria infection by targeting a new site of vulnerability on the parasite
- PMID: 29554083
- PMCID: PMC5893371
- DOI: 10.1038/nm.4512
A human monoclonal antibody prevents malaria infection by targeting a new site of vulnerability on the parasite
Erratum in
-
Author Correction: A human monoclonal antibody prevents malaria infection by targeting a new site of vulnerability on the parasite.Nat Med. 2019 Jan;25(1):188-189. doi: 10.1038/s41591-018-0315-0. Nat Med. 2019. PMID: 30552419
Abstract
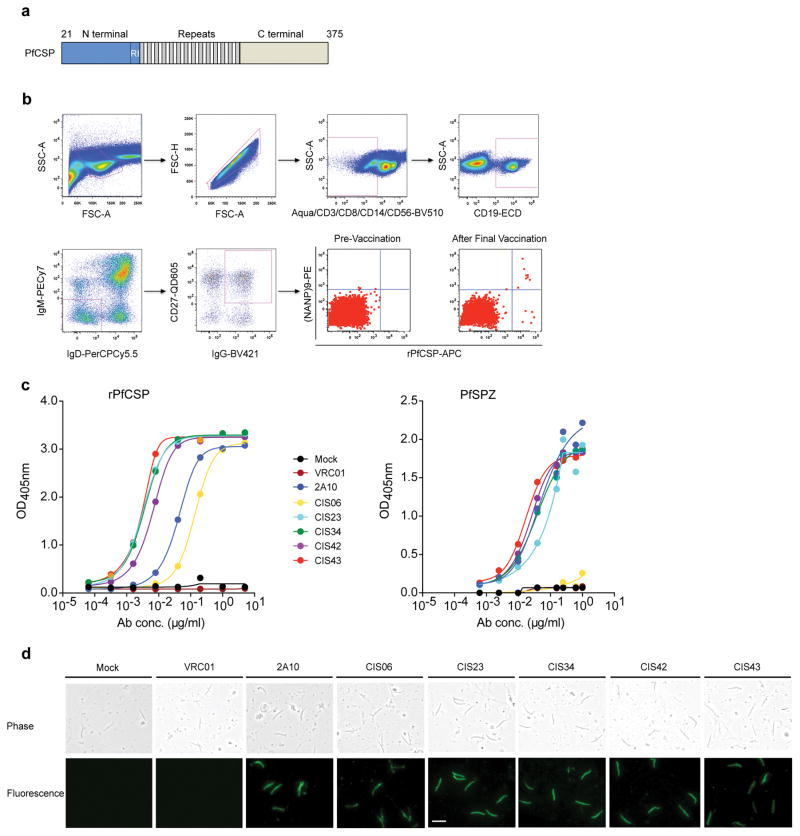
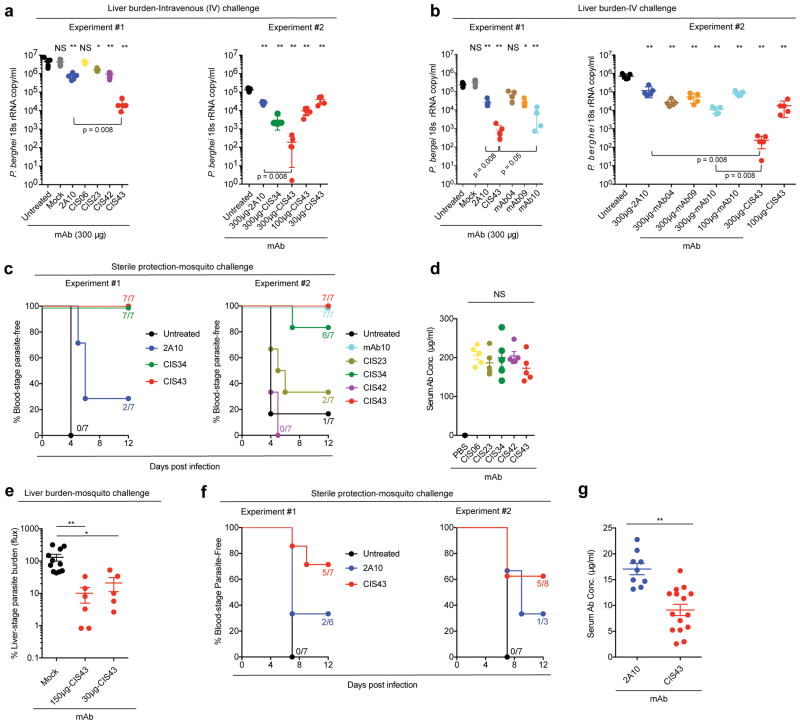
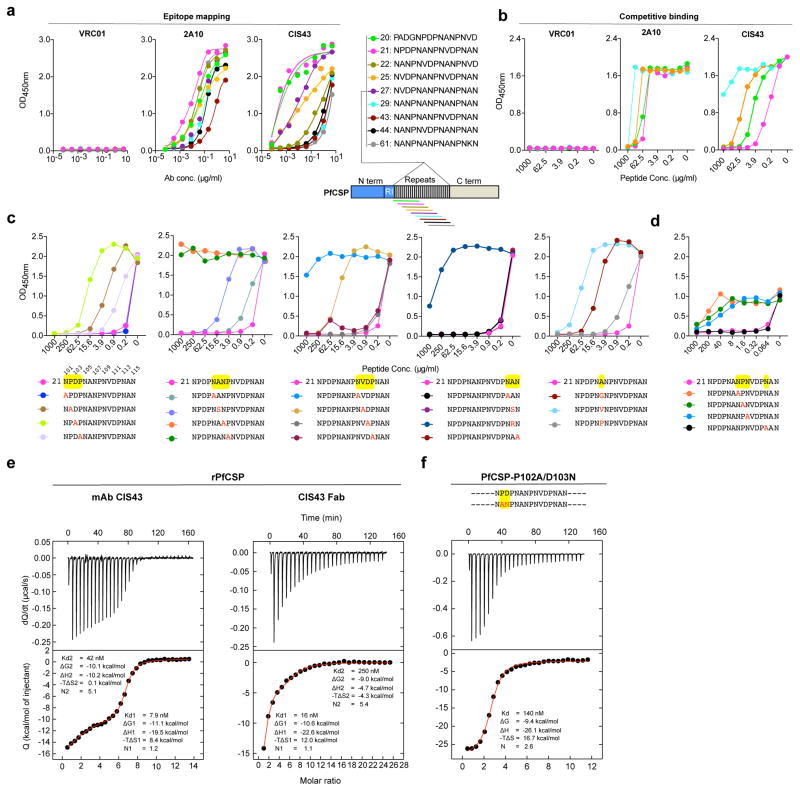
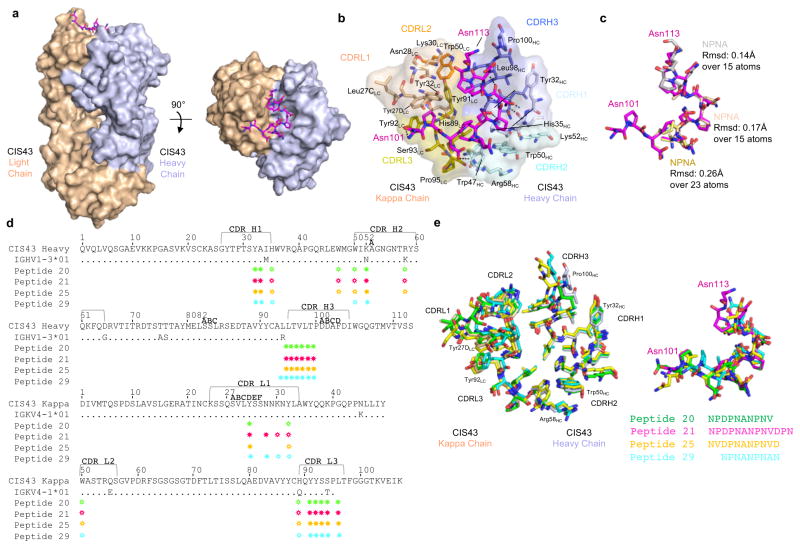
Development of a highly effective vaccine or antibodies for the prevention and ultimately elimination of malaria is urgently needed. Here we report the isolation of a number of human monoclonal antibodies directed against the Plasmodium falciparum (Pf) circumsporozoite protein (PfCSP) from several subjects immunized with an attenuated Pf whole-sporozoite (SPZ) vaccine (Sanaria PfSPZ Vaccine). Passive transfer of one of these antibodies, monoclonal antibody CIS43, conferred high-level, sterile protection in two different mouse models of malaria infection. The affinity and stoichiometry of CIS43 binding to PfCSP indicate that there are two sequential multivalent binding events encompassing the repeat domain. The first binding event is to a unique 'junctional' epitope positioned between the N terminus and the central repeat domain of PfCSP. Moreover, CIS43 prevented proteolytic cleavage of PfCSP on PfSPZ. Analysis of crystal structures of the CIS43 antigen-binding fragment in complex with the junctional epitope determined the molecular interactions of binding, revealed the epitope's conformational flexibility and defined Asn-Pro-Asn (NPN) as the structural repeat motif. The demonstration that CIS43 is highly effective for passive prevention of malaria has potential application for use in travelers, military personnel and elimination campaigns and identifies a new and conserved site of vulnerability on PfCSP for next-generation rational vaccine design.
Conflict of interest statement
N.K.C., S.C., B.K.L.S., and S.L.H. are salaried employees of Sanaria Inc., the developer and owner of PfSPZ Vaccine and the sponsor of the clinical trials. In addition, S.L.H. and B.K.L.S. have a financial interest in Sanaria Inc. All other authors declare no competing financial interests.
Figures
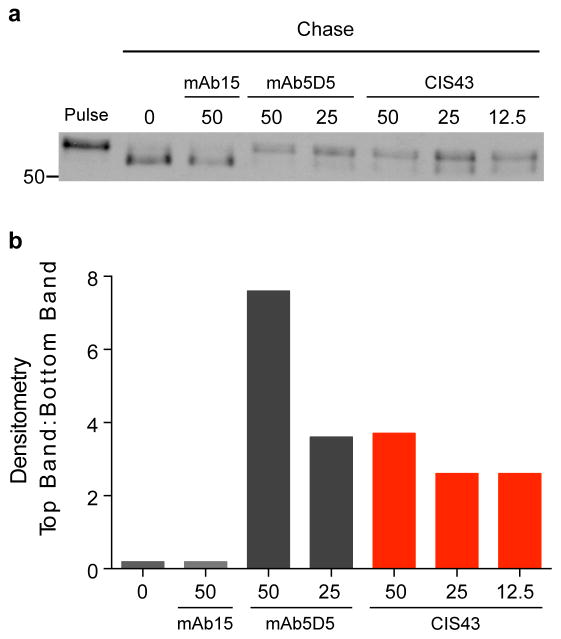
Comment in
-
Peek-Peak-Pique: Repeating Motifs of Subtle Variance Are Targets for Potent Malaria Antibodies.Immunity. 2018 May 15;48(5):851-854. doi: 10.1016/j.immuni.2018.04.037. Immunity. 2018. PMID: 29768173
Comment on
-
A public antibody lineage that potently inhibits malaria infection through dual binding to the circumsporozoite protein.Nat Med. 2018 May;24(4):401-407. doi: 10.1038/nm.4513. Epub 2018 Mar 19. Nat Med. 2018. PMID: 29554084 Free PMC article.
-
A new site of attack for a malaria vaccine.Nat Med. 2018 Apr 10;24(4):382-383. doi: 10.1038/nm.4533. Nat Med. 2018. PMID: 29634679 No abstract available.
References
-
- Dame JB, et al. Structure of the gene encoding the immunodominant surface antigen on the sporozoite of the human malaria parasite Plasmodium falciparum. Science. 1984;225:593–599. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
