

Activity and fidelity of human DNA polymerase α depend on primer structure
- PMID: 29555682
- PMCID: PMC5936803
- DOI: 10.1074/jbc.RA117.001074
Activity and fidelity of human DNA polymerase α depend on primer structure
Abstract
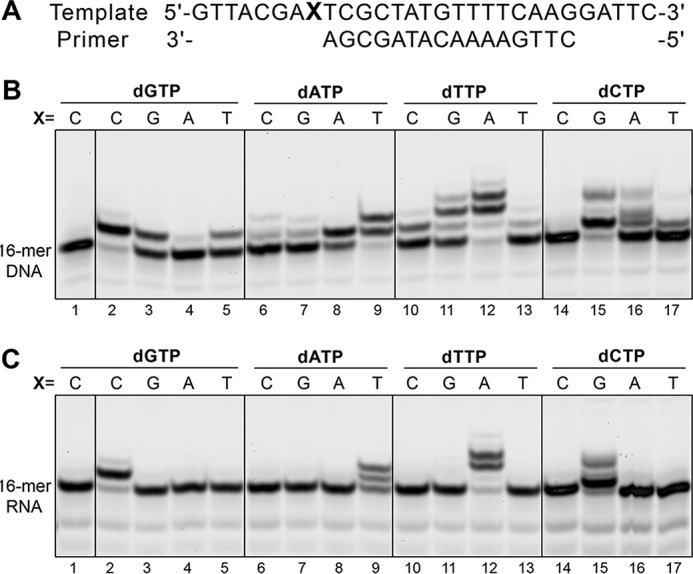
DNA polymerase α (Polα) plays an important role in genome replication. In a complex with primase, Polα synthesizes chimeric RNA-DNA primers necessary for replication of both chromosomal DNA strands. During RNA primer extension with deoxyribonucleotides, Polα needs to use double-stranded helical substrates having different structures. Here, we provide a detailed structure-function analysis of human Polα's interaction with dNTPs and DNA templates primed with RNA, chimeric RNA-DNA, or DNA. We report the crystal structures of two ternary complexes of the Polα catalytic domain containing dCTP, a DNA template, and either a DNA or an RNA primer. Unexpectedly, in the ternary complex with a DNA:DNA duplex and dCTP, the "fingers" subdomain of Polα is in the open conformation. Polα induces conformational changes in the DNA and hybrid duplexes to produce the universal double helix form. Pre-steady-state kinetic studies indicated for both duplex types that chemical catalysis rather than product release is the rate-limiting step. Moreover, human Polα extended DNA primers with higher efficiency but lower processivity than it did with RNA and chimeric primers. Polα has a substantial propensity to make errors during DNA synthesis, and we observed that its fidelity depends on the type of sugar at the primer 3'-end. A detailed structural comparison of Polα with other replicative DNA polymerases disclosed common features and some differences, which may reflect the specialization of each polymerase in genome replication.
Keywords: DNA polymerase; DNA replication; DNA-protein interaction; Polα; Polδ; Polε; RNA; chimeric RNA-DNA primer; conformational change; crystal structure; human; pre-steady-state kinetics.
© 2018 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures













References
-
- Núñez-Ramírez R., Klinge S., Sauguet L., Melero R., Recuero-Checa M. A., Kilkenny M., Perera R. L., García-Alvarez B., Hall R. J., Nogales E., Pellegrini L., and Llorca O. (2011) Flexible tethering of primase and DNA Pol α in the eukaryotic primosome. Nucleic Acids Res. 39, 8187–8199 10.1093/nar/gkr534 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
