

Depletion of Glucose Activates Catabolite Repression during Pneumonic Plague
- PMID: 29555700
- PMCID: PMC5952388
- DOI: 10.1128/JB.00737-17
Depletion of Glucose Activates Catabolite Repression during Pneumonic Plague
Abstract
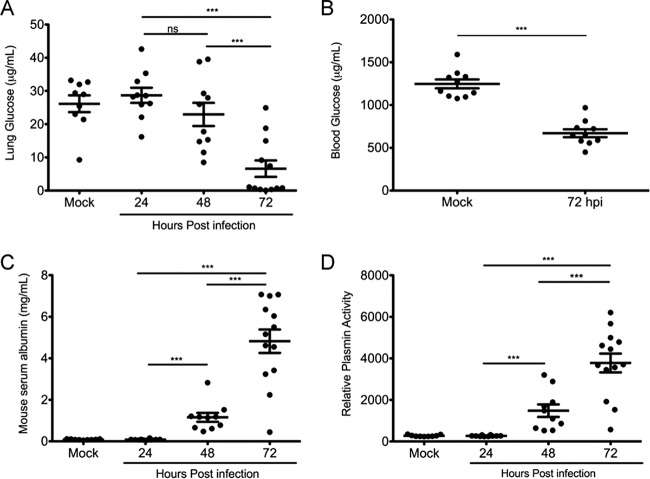
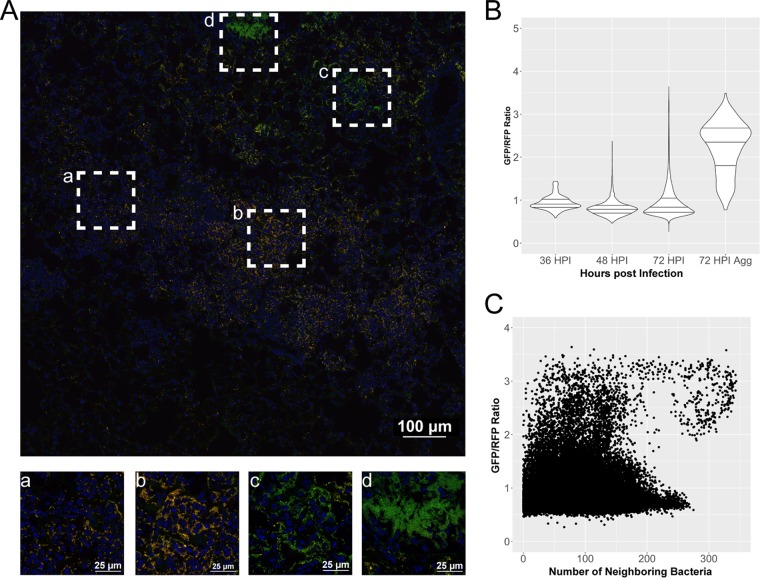
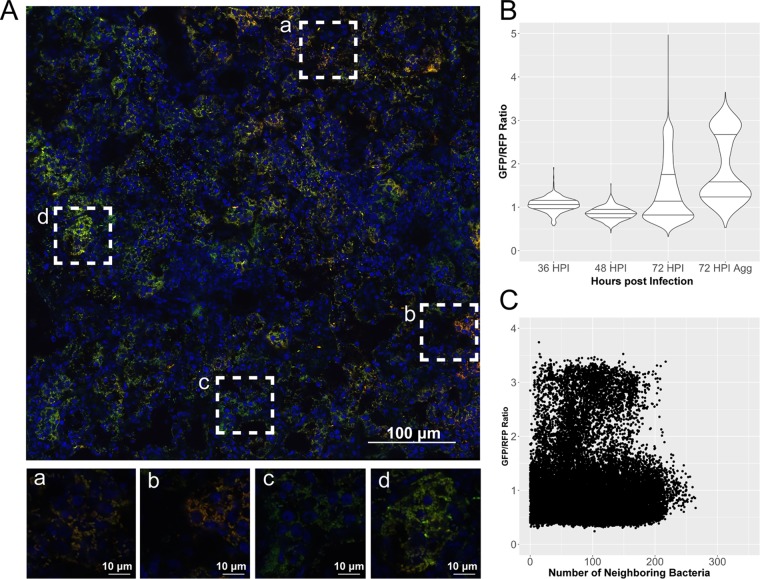
Bacterial pathogenesis depends on changes in metabolic and virulence gene expression in response to changes within a pathogen's environment. The plague-causing pathogen, Yersinia pestis, requires expression of the gene encoding the Pla protease for progression of pneumonic plague. The catabolite repressor protein Crp, a global transcriptional regulator, may serve as the activator of pla in response to changes within the lungs as disease progresses. By using gene reporter fusions, the spatial and temporal activation of the crp and pla promoters was measured in a mouse model of pneumonic plague. In the lungs, crp was highly expressed in bacteria found within large aggregates resembling biofilms, while pla expression increased over time independent of the aggregated state. Increased expression of crp and pla correlated with a reduction in lung glucose levels. Deletion of the glucose-specific phosphotransferase system EIIBC (PtsG) of Y. pestis rescued glucose levels in the lungs, resulting in reduced expression of both crp and pla We propose that activation of pla expression during pneumonic plague is driven by an increase of both Crp and cAMP levels following consumption of available glucose in the lungs by Y. pestis Thus, Crp operates as a sensor linking the nutritional environment of the host to regulation of virulence gene expression.IMPORTANCE Using Yersinia pestis as a model for pneumonia, we discovered that glucose is rapidly consumed, leading to a catabolite-repressive environment in the lungs. As a result, expression of the gene encoding the plasminogen activator protease, a target of the catabolite repressor protein required for Y. pestis pathogenesis, is activated. Interestingly, expression of the catabolite repressor protein itself was also increased in the absence of glucose but only in biofilms. The data presented here demonstrate how a bacterial pathogen senses changes within its environment to coordinate metabolism and virulence gene expression.
Keywords: Crp; Pla; PtsG; Yersinia pestis; catabolite repression; glucose; plague.
Copyright © 2018 American Society for Microbiology.
Figures
References
-
- Chauvaux S, Rosso ML, Frangeul L, Lacroix C, Labarre L, Schiavo A, Marceau M, Dillies MA, Foulon J, Coppee JY, Medigue C, Simonet M, Carniel E. 2007. Transcriptome analysis of Yersinia pestis in human plasma: an approach for discovering bacterial genes involved in septicaemic plague. Microbiology 153:3112–3124. doi: 10.1099/mic.0.2007/006213-0. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
